Živočichové a prostředí
Ekologické systémy
Ekologie bývá nejčastěji definována jako věda o vzájemných vztazích mezi organismy a jejich prostředím. Její název je odvozen z řeckého oikos = dům, obydlí. Předmětem ekologických studií jsou ty systémy, v nichž je alespoň jeden z prvků živý organismus. Ekologie se zabývá třemi stupni: jednotlivým organismem, populací (složenou z jedinců téhož druhu) a společenstvem (složeným z většího či menšího počtu populací). Ekologii na úrovni jedince studuje autekologie, ekologií populací se zabývá demekologie a ekologií společenstev synekologie. Společenstvo neboli biocenóza je tvořeno živočichy - zoocenóza a rostlinami – fytocenóza. Zoocenózy dále dělíme na entomocenózy, ichtyocenózy, ornitocenózy apod. Dílčí soubory populací resp. příbuzných organismů podobné tělesné stavby, obývající totéž prostředí (např. kopytníci afrických savan, norující hlodavci stepí či pouští), označujeme jako taxocenózy (termín zdůrazňuje společnou taxonomickou příslušnost) nebo synuzie (zdůrazněno funkční zapojení). Stále více se prosazuje termín gilda (z angl. guild = cech), kde pojítkem mezi ekologicky spjatými druhy je podobná potrava - např. zimní hejnka sýkor. Každý jedinec, druh nebo společenstvo organismů žije v přírodě v určitém prostředí, které mu poskytuje vhodné podmínky pro život. Toto místo se nazývá biotop (stanoviště, habitat). Biotopy, na nichž byl výskyt daného druhu konkrétně prokázán, se nazývají naleziště (lokality). Živé systémy (živočichové i rostliny) spolu se systémem prostředí (ekotop - prostředí definované souborem abiotických činitelů) tvoří společně dynamické ekologické soustavy – ekosystémy. Rozsáhlé soubory podobných suchozemských ekosystémů nazýváme biomy. Ekosystémy jsou základní jednotky celého biologického systému Země – biosféry. Ekosystém můžeme tedy definovat jako otevřený, dynamicky se vyvíjející živý systém, v němž dochází ke koloběhu látek a v němž probíhá tok energie a výměna genetických informací. V ekosystémech působí autoregulace nastolující dynamickou rovnováhu a snižující entropii. Ekosystémem je např. tundra, rybník, les nebo pole, také to však může být akvárium nebo zkumavka s mikroorganismy.
Ekologické faktory a ekologická valence
Za ekologické faktory považujeme všechny činitele, které nějakým způsobem působí na živočichy, a tím je také ovlivňují. Podle jejich původu je dělíme na abiotické a biotické faktory. Mezi první počítáme veškeré fyzikální a chemické faktory ovzduší, půdy a vodního prostředí, k činitelům biotickým vnitrodruhové a mezidruhové vztahy. Antropogenní faktory jsou dány lidskou činností v ekosystému.
Každý živočišný druh obsazuje v ekosystému určitý prostor a současně je do něho zapojen. Říkáme, že zabírá určitou ekologickou niku (nika = výklenek v antické architektuře pro sochu bohyně Níké). Ekologická nika je tedy funkční začlenění druhu do struktury a funkcí ekosystému. Jinak ji lze definovat jako soubor nároků určitého druhu na podmínky prostředí. To znamená, že organismy kteréhokoliv druhu mohou přežívat, růst, rozmnožovat se a zachovávat životaschopnou populaci jen v rozmezí jistých hranic podmínek, které působí jako rozměry niky. Nika může být potravní (trofická), prostorová, časová (např. při lovu členovců ve vzduchu střídání "denní a noční směny" - vlaštovky a netopýři) apod. Jeden druh nemůže obsadit dvě niky a naopak jedna nika nemůže být obsazena dvěma druhy.
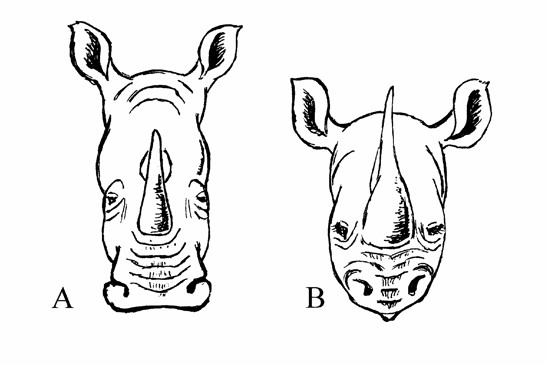 Obr. Využití různých potravních nik dvěma blízce příbuznými sympatrickými druhy. Nosorožec tuponosý (A) má rovné široké pysky uzpůsobené k pastvě – škubání trávy (říká se mu také nosorožec širokohubý). Nosorožec dvourohý (B) má na horním pysku jakýsi „prstík“ usnadňující okusování listí z větviček keřů. Angličtina má pro tento způsob výživy název „browsing“ (JK podle Veselovského, 1974).
Obr. Využití různých potravních nik dvěma blízce příbuznými sympatrickými druhy. Nosorožec tuponosý (A) má rovné široké pysky uzpůsobené k pastvě – škubání trávy (říká se mu také nosorožec širokohubý). Nosorožec dvourohý (B) má na horním pysku jakýsi „prstík“ usnadňující okusování listí z větviček keřů. Angličtina má pro tento způsob výživy název „browsing“ (JK podle Veselovského, 1974).
Tolerancí označujeme schopnost živočichů snášet určité rozpětí libovolného faktoru. Rozpětí tolerance (hranice mezi minimální a maximální mírou působení) se nazývá ekologická valence druhu. Druhy s valencí úzkou označujeme jako stenovalentní, naopak druhy se širokou valencí jsou euryvalentní. Živočichové jsou rovněž různě citliví na celkový charakter stanoviště a obývaného prostředí. Druhy, které obývají různá stanoviště, např. žijí na polích, loukách i v lesích, označujeme jako eurytopní, druhy stanovištně vyhraněné jsou stenotopní. Euryekní druhy se vyskytují v nejrůznějších podmínkách, zatímco druhy stenoekní v podmínkách velmi specifických. U nás se např. s pěnicí černohlavou nebo pěnkavou setkáme od městských zahrad a lužních lesů nížin až po "pralesní" horské smrčiny u horní hranice lesa, kdežto s rákosníkem velkým pouze v rákosinách.
 Obr. Levhart skvrnitý je šelmou s velkou ekologickou valencí a obrovským areálem. Z tropických savan a deštných lesů Afriky a Indonésie zasahuje až do pouští Arábie a lesů ruského Dálného východu, kde v zimě sněží a silně mrzne (Foto OS, Zoo Jihlava).
Obr. Levhart skvrnitý je šelmou s velkou ekologickou valencí a obrovským areálem. Z tropických savan a deštných lesů Afriky a Indonésie zasahuje až do pouští Arábie a lesů ruského Dálného východu, kde v zimě sněží a silně mrzne (Foto OS, Zoo Jihlava).
Ekologické faktory, které působí v rozsahu mezních hodnot, jsou pro přežití jedinců zvláště kritické a nazýváme je mezní neboli limitující faktory. Ty vtiskují konečný ráz různým, zvláště extrémním prostředím, protože mají rozhodující význam pro výběr druhů, hlavně málo pohyblivých nebo stálých, které jsou na daný typ prostředí velmi těsně vázány. Jestliže na organismus působí komplexně celý soubor faktorů, pak platí, že pro přežití a zdárný vývoj jedince musí být hodnoty naprosto všech v rozmezí ekologické valence pro příslušný druh. To znamená, že úspěšnost organismu v daném prostředí je limitována tím faktorem, jehož hodnota je mu nejméně příznivá.Znalost ekologické valence jednotlivých druhů lze využít i k tzv. bioindikaci. Druhy, které jsou citlivé k určitému faktoru a signalizují jeho působení, nazýváme ekologické indikátory. Ty upozorňují na míru antropogenních vlivů, jsou ukazateli přirozených změn nebo indikují vlastnosti abiotického prostředí. Negativní změny jsou obvykle indikovány ubýváním nebo vymizením sledovaných druhů.
Potravní ekologie
Potrava patří mezi rozhodující ekologické faktory. Živočichy řadíme mezi heterotrofní organismy, získávající energii rozkladem organických látek vytvořených autotrofy, které jsou také rozhodujícím zdrojem výživy. Tyto organické látky ve svých tělech různě přetvářejí a opět postupně přeměňují v látky anorganické.
Podle šířky potravního spektra můžeme živočichy zařadit mezi monofágy (živí se jen jediným typem potravy), oligofágy (živí se několika typy potravy) nebo polyfágy (spotřebovávají mnoho typů potravy). Nesmíme si ovšem myslet, že polyfágové a oligofágové přijímají jako přijatelnou potravu všechno a bez rozdílu. Právě naopak, téměř vždy je tu zřejmý jistý stupeň výběru, který je definován srovnáním potravního spektra s dostupností jednotlivých složek potravy.
Biofágové konzumují živá těla organismů nebo jejich části. Dělíme je např. na fytofágy, kteří se živí rostlinami, zoofágy, kteří požírají živočišné organismy, dále druhy mykofágní (požírající houby) a bakteriofágní (bakterie). Fytofágové mohou být různě specializovaní na určitou systematickou skupinu rostlin nebo určitý rostlinný orgán. Herbivoři jsou zpravidla větších rozměrů a konzumují podstatnou část rostliny, naopak fytoparazité (rostlinní cizopasníci) jsou drobní a rostlinu jen poškozují. Parazité bývají vesměs specializovanější na druh potravy i napadenou část rostliny. Zvláštním typem fytoparazitů je minující a hálkotvorný hmyz. Fytofágové mohou být xylofágní (požírají dřevo), fruktivorní (plody), mellivorní (nektar), polenivorní (pyl), fylofágní (listy), granivorní (zrní), graminivorní (trávu), algofágní (řasy), lichenofágní (lišejníky).
Zoofágy dělíme na predátory a parazity. Jednoduše řečeno, predace je spotřebovávání jednoho organismu (kořisti) jiným organismem (predátorem). Při prvním napadení predátorem je kořist živá. Tím je z této kategorie vyloučena detritovorie, což je konzumace mrtvé organické hmoty v různém stupni rozkladu, kterou způsobují saprofágové. Z nich potom nekrofágové konzumují mrtvá těla organismů, zatímco koprofágové se živí trusem živočichů. Zoofágy dělíme např. na myrmekofágy (požírají mravence), ichtyofágy (ryby), hematofágy (krev), ornitofágy (ptáky), entomofágy (hmyz) a karnivory (maso). Někdy existují synonymické dvojice názvů, jeden latinského, druhý řeckého původu - např. piscivorní (rybožravý), insektivorní (hmyzožravý). Paraziti (cizopasníci) konzumují části své kořisti (svého hostitele), nikoliv celou kořist. Jejich napadení jsou škodlivá, ale jen zřídka mohou v krátké době přivodit smrt. Cizopasníci se během svého života soustřeďují na jednoho či jen na několik málo jedinců, a proto mezi parazity a jejich hostiteli se často vytváří těsné spojení. Ze specifických způsobů získávání potravy lze jmenovat např. požírání substrátu (kroužkovci) nebo mikrofágii, tj. filtrování (houbovci a další bezobratlí planktonofágové, plameňáci, kachny, žralok obrovský, velryby).
Potravní vztahy v ekosystémech vyjadřuje ekologická potravní pyramida. Vyplývá z ní, že jediným zdrojem organické hmoty ekosystému jsou autotrofní organismy (zelené rostliny a některé bakterie) – producenti. Ti se stávají zdrojem pro heterotrofní organismy - konzumenty (tj. organismy, které vyžadují zdroje v organické, energeticky bohaté formě) a účastní se celého řetězce dějů, ve kterém se každý spotřebitel zdrojů (konzument) stává naopak zdrojem pro dalšího konzumenta. Takový sled několika úrovní postupně se konzumujících organismů nazýváme potravní řetězec. Prvním článkem každého potravního řetězce je tedy autotrofní organismus nazývaný producent (P). Od producenta vede řetězec přes fytofágy a bakteriofágy, tj. konzumenty 1. řádu (primární konzumenti, K1) k několika úrovním zoofágů, konzumentů vyšších řádů (sekundární konzumenti, K2, K3 atd.). Na bázi potravní pyramidy jsou tedy producenti, na špičce vrcholoví predátoři. V případě predátorů se obvykle na dané trofické úrovni zvětšuje velikost těla a počet jedinců klesá, pokud je konzumentem parazit, velikost těla bývá menší než na předešlé úrovni a jedinců je často více. Mrtvá těla organismů na všech úrovních (P, K1 až Kn) jsou konzumována saprofágy a dekompozitory v tzv. dekompozičním řetězci. Tělesná velikost dekompozitorů bývá velmi malá, avšak dosahují vysoké populační hustoty. Čím je síť potravních řetězců složitější, tím je pevnější a ekosystém je stabilnější.
Na každé potravní úrovni se část energie spotřebovává ke krytí životních funkcí organismu a část energie se uvolňuje ve formě tepla. Živočichové na příslušné potravní úrovni využívají jen asi 10 % látek a energie z úrovně předcházející.
Velikost populace a rozmístění jedinců v populaci
Živočichové v přírodě nežijí osamoceně, nýbrž vytvářejí populace, které můžeme definovat jako soubor jedinců téhož druhu na určitém území. Každá populace se vyznačuje svojí velikostí, která je dána počtem jedinců na jednotku plochy či prostoru. Ukazatelem kvantitativního zastoupení živočišné populace v prostředí je abundance (početnost). Celkový počet jedinců, tzv. absolutní abundance, se zjišťuje spíše ve výjimečných případech. Je to možné například u velkých savců nebo v hnízdních koloniích ptáků, kde lze jedince snadno pozorovat a počítat nebo odlovit. Častěji určujeme abundanci relativní, což je počet odchycených nebo pozorovaných jedinců určitou metodou. Pokud je možné abundanci vyjádřit ve vztahu k určité ploše nebo prostoru (m2, m3, ha), hovoříme většinou o hustotě (denzitě) populace. Studium početnosti populací má značný význam zejména u vymírajících druhů, u populací druhů využívaných člověkem a u druhů, které při zvýšených hustotách působí škody.
Vzhledem k rozdílné velikosti jedinců v rámci jednotlivých druhů se může hustota přepočítat na hmotnost živé hmoty (organismů) – biomasu. Velikost biomasy vyjadřujeme nejčastěji hmotností jedinců a je ji možno opět uvádět v absolutních hodnotách celé populace, účelnější a využitelnější jsou však údaje přepočítané na jednotky plochy nebo objemu.
Při zjišťování početnosti i biomasy musíme vycházet z předpokladu nerovnoměrného rozmístění jedinců v prostoru. Proto je nutné pozorování vícekrát zopakovat a vypočítat průměrnou hodnotu. Hodnoty těchto ukazatelů se také různě mění v průběhu roku i během delších časových období. O tom již více pojednává další text.
Rozmístění jedinců v populaci - rozptyl (disperze), bývá u živočišných druhů zpravidla nerovnoměrné a nepravidelné, často se snahou shlukovat se. Disperze také informuje o umístění jedinců v obývaném prostoru, proto rozlišujeme lineární disperzi (např. u některých druhů obývajících břehové porosty tekoucích vod), plošnou disperzi (např. obratlovci obývající zemský povrch nebo žijící těsně pod ním) a prostorová disperze (např. létaví a stromoví ptáci, šplhaví savci a vodní obratlovci, obývající různé vodní vrstvy).
Populační dynamika živočichů
Populace je živý systém, v němž se projevují biologické vlastnosti jedinců, ale i biologické vlastnosti celé skupiny. Populační dynamika živočichů nám znázorňuje kolísání početnosti populace, ke kterému dochází v různých časových intervalech. Tyto změny bývají způsobeny natalitou, mortalitou a částečně také migralitou jedinců. Natalita (porodnost, množivost) je schopnost populace narůstat, přičemž ji vyjadřujeme počtem nových jedinců za jednotku času. Rozlišujeme maximální (fyziologickou) množivost, která představuje maximální produkci nových jedinců za ideálních vnitřních i vnějších podmínek a realizovanou (ekologickou) množivost, která je výsledkem působení okamžitých podmínek prostředí na populaci. Úbytek jedinců v populaci v důsledku vymírání označujeme jako mortalitu (úmrtnost). Rovněž úmrtnost může být minimální (fyziologická) a realizovaná (ekologická). S úmrtností velmi úzce souvisí i délka života jedince určitého druhu. U řady populací dochází k pravidelným nebo nepravidelným výkyvům populační hustoty. Ke kolísání početnosti může docházet buď během jediného roku, což označujeme jako oscilace nebo kolísání během více let -tzv. fluktuace. V obou případech se jedná o výsledek rozdílu mezi natalitou a mortalitou, ale teprve studium většího počtu oscilací, tzn. fluktuace nám většinou ukáže, k jakým výkyvům populační hustoty skutečně dochází. Některé druhy živočichů mají schopnost se čas od času přemnožovat (hraboši, lumíci, zajíc měnivý). To označujeme jako gradace, při které dochází k extrémnímu vzestupu početnosti populace. U každé gradace rozlišujeme fázi nízké početnosti, tzv. latence, po níž následuje vzestupná fáze zakončená kulminací, tj. gradačním vrcholem. Po překonání maxima dochází k prudkému poklesu populační hustoty (úhyn v důsledku stresu, nemocí) až opět do stadia latence. Na kolísání početnosti populací mají obvykle vliv klimatické podmínky, vzájemné vztahy kořisti a predátorů, parazité, nemoci, zásahy člověka, ale i vnitřní fyziologické a genetické činitele. Cyklické přemnožování je nejvýraznější v jednodušších ekosystémech (tundra, monokultury).
Stěhování a šíření populace
Soubor schopností určitého druhu, které mu umožňují se šířit na vhodná stanoviště uvnitř dosavadního areálu i mimo něj, se nazývá vagilita. Způsoby přemísťování sahají od pasivního přenosu, až po aktivní aktivní pohyb mnoha mobilních živočichů. K popisu některých aspektů pohybu živočišných organismů se používá termínů disperze (rozmístění, rozptyl) a migrace. Podle směru pohybu dále rozlišujeme emigraci (vystěhování) a imigraci (přistěhování). Migrace většinou znamená hromadný směrovaný pohyb většího počtu jedinců nějakého druhu z jednoho místa na jiné. Disperze nejčastěji označuje vzdalování se jedinců od jiných (např. od jejich rodičů či sourozenců), patří sem jak pohyby aktivní (chůze, plavání, létání), tak pohyby pasivní (přenos vodou, větrem či prostřednictvím jiných živočichů). Ke stěhování a šíření dochází v různých fázích vývoje. Rozhodujícími příčinami těchto přesunů jsou klima, potrava, rozmnožování a prostor. Migralita má nesmírný význam pro předávání dědičných informací mezi jednotlivými populacemi. V některých případech také ovlivňuje populační hustotu. Periodické migrace jsou patrně nejlépe prostudovány u ptáků. Velmi známé jsou také tahy ryb. Z jiných obratlovců byly pozorovány různě dlouhé migrace u savců (sobi, pakoně), plazů (mořské želvy) i obojživelníků. U členovců se vyskytují jak aktivní, tak pasivní formy migrality. Drobné druhy hmyzu, pavouků a roztočů mohou být přenášeny větrem nebo prostřednictvím jiných živočichů. K aktivním migracím dochází např. u motýlů (babočka bodláková, monarcha) nebo sarančí. K šíření živočichů dochází i lidskou činností (antropochorní přenos), a to jednak záměrnou introdukcí, jednak neúmyslným zavlékáním organismů. Introdukcí rozumíme zavádění nových živočišných druhů do oblastí, kde se dříve nevyskytovaly, naproti tomu reintrodukce (repatriace) druhu je vysazení do původního areálu, v němž byl druh v minulosti vyhuben nebo kde se jeho stavy drasticky snížily. Invaze jednotlivých druhů mají různý charakter, probíhají různě rychle a zasáhnou různě velké území. V počáteční fázi je invaze pomalá a její rychlost se postupně zvětšuje. Po vyčerpání možností šíření a nastolení rovnováhy s přirozenými antagonisty se invaze zpomaluje až zastavuje.
Vztahy mezi populacemi různých druhů
Mezi různými druhy organismů dochází v přírodě k rozmanitým formám soužití. Žádný živočišný druh není schopen existovat samostatně, izolovaně od ostatních. Jakýkoli současný stav mezi dvěma druhy nebo populacemi je výsledkem dlouhodobého souběžného vývoje, který nazýváme koevoluce. Výsledkem vzájemné interakce je pozitivní nebo negativní ovlivňování alespoň jedné nebo obou zúčastněných populací. Základní typy vztahů mezi populacemi uvádí následující tabulka.
| Název vztahu | Populace | |
|---|---|---|
| A | B | |
| Neutralismus | 0 | 0 |
| Amenzalismus | - | 0 |
| Predace, herbivorie, parazitismus | + | - |
| Komenzalismus | + | 0 |
| Protokooperace, symbióza | + | + |
| Konkurence | - | - |

 Obr. Štírci se mohou šířit na větší vzdálenost pomocí tzv. foreze. Jde o využívání létajícího hmyzu, na který se štírci většinou přichycují pomocí pedipalp. Na obr. jsou štírci přichycení na mouše domácí (Česká republika) a na brouku kovaříkovi (Kostarika) (Foto OS).
Obr. Štírci se mohou šířit na větší vzdálenost pomocí tzv. foreze. Jde o využívání létajícího hmyzu, na který se štírci většinou přichycují pomocí pedipalp. Na obr. jsou štírci přichycení na mouše domácí (Česká republika) a na brouku kovaříkovi (Kostarika) (Foto OS).
Parazitičtí kastrátoři hostitele nezabijí, ale zabrání mu v reprodukci, což je z hlediska evoluce totéž (snížení fitness na nulu). Mikroparazité se v těle hostitele (mnohdy přímo v buňkách) množí a bývá jich velký počet (viry, bakterie, prvoci), kdežto makroparazité v něm svůj počet nezvětšují, jen produkují stadia infekční pro další hostitele (helminti, členovci). Spíše než v buňkách jsou lokalizováni intercelulárně, v tělních dutinách nebo dutých orgánech. Zde je třeba si uvědomit, že parazitismus je ekologický vztah, nikoli systematické zařazení, jak se dříve někdy tradovalo (prvoci, helminti a členovci byli pokládáni za parazity, viry a bakterie nikoli). Parazitem tedy může být nejen virus, bakterie, rostlina či houba, ale i obratlovec (kukačka).
Parazité jsou menší než hostitel a žijí na jeho povrchu (ektoparazité) nebo uvnitř jeho těla (endoparazité), uvnitř nebo vně buněk (intra- nebo extracelulárně). Parazity dělíme na jednohostitelské (monoxenní) nebo vícehostitelské (heteroxenní). U definitivního (finálního) hostitele probíhá pohlavní fáze rozmnožování parazita resp. v něm žije dospělec, u mezihostitele množení neprobíhá nebo je nepohlavní. V paratenickém (transportním) hostiteli vývoj nepokračuje a dochází k hromadění parazitů. Přenašeč se označuje též vektor. Hyperparazit je parazitem jiného parazita. Parazité jsou obecně specializovanější než predátoři a herbivoři. Navíc jde v případě parazitismu v některých případech o vztah tří nebo více populací. Do vývojových cyklů totiž vstupují populace mezihostitelů, přenašečů (vektorů) nebo paratenických hostitelů. Parazitismus může být trvalý (stacionární) nebo dočasný (temporární). Dochází k němu zcela pravidelně (obligátní parazitismus) nebo jen příležitostně (fakultativní parazitismus). Z hlediska hostitelské specifity jsou parazité euryxenní (např. toxoplazma) nebo stenoxenní (např. "všenky"). Sociální parazité zneužívají sociálního chování jiných jedinců k jednostrannému zvyšování vlastní fitness; existují hlavně u sociálního blanokřídlého hmyzu, zejména mravenců, ale známe je i z lidské společnosti. Formou sociálního parazitismu je hnízdní parazitismus, známý zvláště u ptáků. Může být vnitrodruhový (některé kachny) nebo mezidruhový (kukačky), fakultativní nebo obligátní.
Předpokládá se, že většina jedinců i druhů organismů je parazitických. Parazité významně ovlivňují nejrůznějšími mechanismy imunitní, endokrinní a nervový systém hostitele. Někteří účinně manipulují chování svých mezihostitelů tak, aby se zvýšila pravděpodobnost jejich pozření definitivním hostitelem. Tak např. myši infikované toxoplazmózou ztrácejí strach z pachu koček, plži jantarky napadení motolicí Leucochloridium vylézají za dne na vyvýšená místa, kde je snáze uvidí a sezobne pták, a konečně mravenci napadení metacerkáriemi motolice kopinaté vylézají na vršky trav, kde jsou spaseni přežvýkavci. Složité vztahy mezi parazity a hostiteli se formovaly během milionů let společného vývoje (koevoluce, kospeciace). Adaptace k parazitismu může vést k extrémní změně morfologie, obvykle ve smyslu zjednodušení stavby těla (rybomorky, jazyčnatky, červok), ztrátě některých orgánů (trávicí trakt, křídla) nebo naopak vývoji nových struktur (přísavky, háčky, bodce aj.). Mohou vznikat komplikované vývojové cykly a značná nadprodukce potomstva.
Populační ekologie parazitů je ovlivněna skutečností, že hostitelé jsou časově a prostorově ohraničení a vzájemně izolovaní a že parazité přílišnou exploatací mohou zničit hostitele, tj. své životní prostředí. Evolučně tak proti sobě působí individuální selekce maximalizující fitness jedince a selekce skupinová maximalizující celkový počet infekčních stadií populace během celé své existence. V dlouhodobé perspektivě převládnou ty druhy, u nichž se vytvořily mechanismy omezující individuální selekci a posilující selekci skupinovou. Takovým omezujícím mechanismem je např. nepohlavní množení (produkce klonů nekonkurujících rodičovské generaci). Izolovanost vede k častému výskytu hermafroditismu. U gonochoristů dochází často k příbuzenské plemenitbě mezi sourozenci a následně k posunu poměru pohlaví ve prospěch samic. Pro populace parazitů je charakteristická jejich shlukovitost (agregovanost) resp. asymetrické rozložení. V závislosti na počtu parazitů v hostiteli se projevuje jejich fenotypová plasticita (různá velikost, rychlost dospívání, plodnost). Parazité různých druhů v hostiteli mohou tvořit strukturovaná společenstva a mohou na sebe přímo či nepřímo negativně nebo pozitivně působit (např. vrtejši a hlístice ve střevě ryb). Navzdory mnohdy obrovské nadprodukci potomstva jsou parazité spíše K-stratégy než r-stratégy, neboť nemaximalizují rychlost, nýbrž efektivnost svého množení. Parazité ovlivňují populační cykly hostitelů a lokální biodiverzitu (i pozitivně), mění fenotyp hostitelů, mění (i zvyšují) produktivitu ekosystémů, ovlivňují areály hostitelů, zvyšují vymírání druhů, ale i speciaci vytvářením reprodukčních bariér. Z uvedeného vyplývá, že interakce parazitů a hostitelů jsou nesmírně komplexní a nelze je zjednodušit na konstatování, že parazité jsou "škůdci" - to neplatí přinejmenším v přírodních ekosystémech.
Velmi rozšířeným vztahem je komenzalismus (doslova soustolovnictví, z lat. mensa = stůl), kdy jedna populace využívá jinou bez jejího poškozování. Nejčastěji jde o závislost potravní (klubáci a volavky provázející africké kopytníky) nebo prostorovou.
 Obr. Nejznámějším příkladem komenzalismu mezi savci a ptáky je vztah mezi africkými kopytníky a klubáky nebo volavkami rusohlavými. Mohli bychom sem zařadit i toto využívání laní v zoologické zahradě strakami (Foto IL).
Obr. Nejznámějším příkladem komenzalismu mezi savci a ptáky je vztah mezi africkými kopytníky a klubáky nebo volavkami rusohlavými. Mohli bychom sem zařadit i toto využívání laní v zoologické zahradě strakami (Foto IL).
Potravní parazitismus (včelomorka) resp. kleptoparazitismus (chaluhy, fregatky) znamená okrádání o potravu. Často se setkáváme s jevem, kdy drobní živočichové vyhledávají blízkost velkého druhu z důvodu větší bezpečnosti, tzv. parekie (ryby lodivodi a žralok). Pokud žijí menší živočichové přímo v hnízdech ptáků, norách savců (nidikolní členovci aj.) nebo v mraveništích (myrmekofilie), jde o tzv. synekii. V případě epiekie se usídlují jedinci jednoho druhu na povrchu těla druhu jiného (štítovec lodivod – žralok), u entekie dokonce uvnitř jeho těla.
Nejrůznější případy oboustranně kladného ovlivňování dvou populací jsou velmi časté. Oboustranně pozitivní soužití může být přitom prospěšné pro populaci jako celek i pro každého jedince nebo pouze pro populaci, i když jedinci jsou likvidováni. Interakce se může realizovat občasným stykem jedinců i trvalým povrchovým nebo vnitřním soužitím. Jednodušší formou pozitivního ovlivňování je protokooperace. Je to vztah sice vzájemně prospěšný, ale není závazný (zimní smíšená hejna sýkor). V průběhu evoluce se postupně vyvinuly svazky závazné, které nazýváme symbióza (mutualismus). Typickým příkladem je symbiotický vztah mezi býložravci a nálevníky v trávicím traktu. V anglosaské literatuře bývá symbióza chápána šířeji, jako jakékoli těsné soužití přinášející prospěch symbiontovi, a tedy zahrnující nejen mutualismus, ale i komenzalismus a parazitismus.
 Obr. Soužití korálových ryb klaunů (Amphiprion spp.) se sasankami vešlo do učebnic jako typický příklad symbiózy. S nárůstem informací o těchto vztazích se ukazuje, že skutečnost je poněkud komplikovanější (Foto JK).
Obr. Soužití korálových ryb klaunů (Amphiprion spp.) se sasankami vešlo do učebnic jako typický příklad symbiózy. S nárůstem informací o těchto vztazích se ukazuje, že skutečnost je poněkud komplikovanější (Foto JK).
Dalším typem vztahu je konkurence (kompetice), kdy populace se vzájemně negativně ovlivňují. Konkurence je tedy typ vztahu mezi jedinci (populacemi) vyvolaného společnou potřebou zdroje, jenž se vyskytuje v omezeném množství, a směřujícího ke snížení možnosti přežití, růstu a reprodukce soutěžících jedinců (populací). Faktor, který je předmětem konkurence, obvykle určuje horní limit početnosti jedinců. Konkurence vede k vytěsnění “slabších” druhů (potkan a krysa obecná, dingo a vakovlk), ale během koevoluce i k diferenciaci a soužití více druhů ve stejném prostoru v mírně odlišných nikách (sýkory, rákosníci). Přispívá tak ke speciaci.
Životní strategie
Z tzv. teorie ostrovní biogeografie (viz kap. Ochrana přírody) vychází koncepce r-strategie a K-strategie. Tzv. r-stratégové jsou druhy, které mají rychlé rozmnožování a velkou populační dynamiku. Jsou krátkověké a jejich populační hustota prodělává velké výkyvy. K-stratégové mají naopak pomalé rozmnožování a malou populační dynamiku, jsou početně poměrně stálí a dlouhověcí. K prvému typu patří např. tzv. gradační druhy drobných hlodavců (hraboš polní, lumíci) nebo mnohé naše ryby. Ke druhému typu náleží např. lidoopi, sloni, velké šelmy a dravci. Oba typy ovšem představují krajní varianty s řadou přechodných strategií. Zkratky se vztahují k označení hodnot v rovnicích, které matematicky popisují příslušné ekologické vztahy a populační růst. Z botaniky pochází poněkud jiné členění, kde např. C-stratégové (z angl. competition = soutěž) žijí ve stálém a "bohatém" prostředí, kde však probíhá ostrá konkurence mezi množstvím druhů. Patří sem např. živočichové tropických deštných lesů a korálových útesů. S-stratégové (od slova stres) jsou citliví ke stresu z narušování, ale tolerantní k nedostatku. Obývají stálá, ale "chudá" prostředí jako jeskyně (často slepé a nepigmentované druhy, např. macarát; tzv. troglobionti); dali by se sem zařadit i praví hibernátoři.
 Obr. Primáti (na snímku pavián čakma) patří ke K-stratégům, tj. savcům s malým počtem mláďat, o která dlouhou dobu pečují (Foto LP).
Obr. Primáti (na snímku pavián čakma) patří ke K-stratégům, tj. savcům s malým počtem mláďat, o která dlouhou dobu pečují (Foto LP).
Dlouhodobý vývoj biocenóz a typy biocenóz
Pod pojmem biocenóza rozumíme soubor populací všech organismů obývajících určité území, které je vymezené souborem abiotických faktorů. Jde o zákonité seskupení vzniklé v čase i prostoru v závislosti na ekotopu a geografických podmínkách. K charakteristickým znakům každé biocenózy patří její stálost (stabilita), nezávislost (autarkie) a autoregulace. Dlouhodobý zákonitý proces vývoje biocenózy se nazývá ekologická sukcese. V jednotlivých fázích jejího vývoje se tvoří sukcesní stadia s charakteristickou vegetací a živočišstvem, která dohromady tvoří sukcesní řadu (sérii) a směřuje od výchozího stavu biocenózy ke konečnému stadiu vývoje biocenózy, kterému říkáme klimax. Klimaxové stadium má největší diverzitu, největší počet mezidruhových vztahů a představuje homeostatický systém (např. v horách ČR klimaxová smrčina). Druhy, které se na neosídleném ekotopu objevují jako první, označujeme pionýrské (např. vrbka úzkolistá po sopečné erupci). Z hlediska zachovalosti (původnosti) rozlišujeme biocenózy přírodní, přirozené a umělé (druhotné). Přírodní biocenózy představují společenstva vzniklá a existující zcela nezávisle na lidské činnosti. S biocenózami tohoto typu se ve střední Evropě dnes téměř nesetkáme, i relativně málo narušená vysokohorská nebo rašeliništní společenstva jsou postižena znečištěním ovzduší, změnami vodního režimu apod. Přirozené biocenózy jsou sice do různé míry ovlivněny lidskou činností, ale svým druhovým složením se blíží přírodnímu stavu v dané oblasti. Mezi takové biocenózy v oblasti střední Evropy patří většinou listnaté, ve vyšších polohách smíšené nebo jehličnaté lesy. Umělá společenstva jsou vytvářena člověkem záměrně (agrocenózy, borové a smrkové monokultury v nížinách ČR), nebo vznikají neřízeně v návaznosti na lidskou činnost (ruderální neboli rumištní biocenózy). Druhotné (sekundární) biocenózy bývají jednotvárnější a druhově chudší než původní (primární) biocenózy (např. sekundární versus primární tropický deštný les). Sekundární les vzniká i přirozeně (sesuvy, sopečné erupce, povodně). Zcela zvláštním typem jsou synantropní biocenózy, která vznikla a udržují se na uměle vytvořených místech, např. v okolí sídlišť. Mnoho lidí si neuvědomuje, že např. louky, většina borů nebo smrkové lesy s výjimkou horských oblastí nejsou v ČR původními společenstvy. Jednotlivé biocenózy nejsou zpravidla od sebe ostře ohraničeny a na místech jejich překrývání vznikají tzv. ekotony neboli přechodná společenstva. Zde se často vyskytuje větší počet druhů než v obou sousedních cenózách, protože do ekotonu pronikají nejen druhy ze sousedních biocenóz, ale také samotný ekoton se vyznačuje určitým počtem specifických druhů. Např. na okraji lesa najdeme větší počet druhů ptáků, než jsou počty v sousedních biocenózách lesa a louky. Podobně v ekologicky vyvážené kulturní krajině může být větší diverzita než v souvislém lese.
Společenstva na území ČR a struktura jejich dílčích společenstev
"Původním" (v holocénu) vegetačním pokryvem na našem území byl převážně les. Dnes jsou v ČR zastoupeny zejména agrocenózy (polní kultury), rozmanité typy lesních společenstev, luční společenstva, synantropní společenstva a biocenózy stojatých a tekoucích vod. V závislosti na nadmořské výšce rozlišujeme (zjednodušeně) lesní vegetační stupeň dubový (do 500 m n.m.), bukový (500-1200 m) popř. jedlobukový, smrkový (1100-1300) a při horní hranici lesa kosodřevinu (kolem 1300 m n.m.). Nad ní jsou ve vysokých horách alpínské louky (hole), skály a sutě, popř. nivální stupeň (sníh a led - Alpy). Smrkový stupeň je vlastně horskou tajgou a alpínský je horskou tundrou. V zaplavovaných nivách řek (aluvia) tvoří původní porost mokřadní olšiny a lužní lesy, členěné na vlhčí tzv. měkký luh (převládají vrby, topoly) a sušší tvrdý luh (jasany, duby). V nížinách a pahorkatinách původně převládaly doubravy a dubohabrové lesy. Přirozené bory nacházíme sporadicky, převážně v Českém masivu. Z hlediska zachování biodiverzity mají nesmírný význam i málo rozšířené typy biocenóz, např. tzv. azonální společenstva - mokřady, rašeliniště, vřesoviště, jeskyně, dále (leso)stepní lokality aj. Příklady typických živočichů: lužní les - moudivláček, tesařík obrovský; doubravy a dubohabřiny - roháč obecný, žluva, brhlík, dlask, šoupálek krátkoprstý; bučiny - mlok skvrnitý, holub doupňák, strakapoud bělohřbetý, puštík bělavý; smrčiny - tetřev, kos horský, křivka; kosodřevina - čečetka zimní, slavík modráček tundrový; alpínský stupeň - svišť, kamzík, linduška horská; mokřady - chřástali, bahňáci, rákosníci; bory - krasec měďák, lelek, skřivan lesní; lesostep - kudlanka nábožná, ještěrka zelená, ťuhýk obecný, pěnice vlašská; rašeliniště - žluťásek borůvkový, tetřívek; skály - zedníček skalní, pěvuška podhorní.
 Obr. Lesní vegetační bukový stupeň zaujímá nižší horské polohy (500 – 1200 m n. m.). Typickými živočichy bučin jsou např. mlok skvrnitý, holub doupňák, strakapoud bělohřbetý nebo lejsek bělokrký (Foto Z. Literáková).
Obr. Lesní vegetační bukový stupeň zaujímá nižší horské polohy (500 – 1200 m n. m.). Typickými živočichy bučin jsou např. mlok skvrnitý, holub doupňák, strakapoud bělohřbetý nebo lejsek bělokrký (Foto Z. Literáková).
Podle prostorového uspořádání můžeme rozdělit biocenózu s jejím biotopem vertikálně a horizontálně. Z hlediska vertikální stratifikace nazýváme jednotlivé vrstvy jako patra, etáže neboli biostrata a dílčí společenstva, která je osídlují, jako stratocenózy (např. stratocenózy korunové, kmenové, křovinné, bylinné, mechové a hrabankové). Prostorově zřetelně vymezené jsou biocenózy i celé ekosystémy stojatých i tekoucích vod. Ve stojatých vodách je možno rozlišit tato dílčí společenstva organismů: Plankton je společenstvo drobných, málo pohyblivých a většinou pasivně unášených organismů ve vodním sloupci, který se dále dělí na fyto- a zooplankton. Nekton je společenstvo aktivně pohyblivých živočichů ve vodním sloupci, kam patří především ryby. Neuston je společenstvo organismů povrchové vodní blanky. Pleuston je společenstvo žijící na hladině a bentos je společenstvo organismů dna nádrže. Tekoucí vody jsou členěny na čtyři rybí pásma, na základě výskytu tzv. vůdčích druhů ryb, a to směrem od pramene k ústí na pásmo pstruhové, lipanové, parmové a cejnové. Soubor půdních organismů se označuje jako edafon a jeho živočišná složka zooedafon. Zástupci zooedafonu rozmělňují a rozrušují organickou hmotu, provzdušňují a převrstvují půdu a podílejí se na tvorbě humusu. Podle výskytu je dělíme na epigeické druhy (žijící na povrchu půdy) a hypogeické druhy, které se vyskytují pod zemí.