Evoluční biologie
Biologická evoluce je dlouhodobý, samostatně probíhající proces, v jehož průběhu vznikají či jednorázově vznikly ze systémů neživých systémy živé, a tyto živé systémy se následně vyvíjejí a diverzifikují. Evoluční biologie studuje vlastnosti procesu biologické evoluce a jeho jednotlivé konkrétní mechanismy. Věnuje se vlastnímu historickému průběhu evolučních procesů v konkrétním prostoru a čase, tedy především vzniku účelných znaků působením přirozeného výběru.
Historie evolučních názorů
V rámci biologie a obecně přírodních věd vznikla řada evolučních teorií zabývajících se vznikem života a jeho evolucí. Tradičně se dělí historie evoluční biologie na tři etapy: na období předdarwinovské, období klasického darwinismu a období neodarwinismu. Dnes se nejčastěji pojmem evoluční teorie rozumí koncepce navazující na Darwina a Wallaceho, v nichž klíčovou roli hraje hromadění nahodilých změn a přírodní výběr.
Období předdarwinovské
První formulace evolučních názorů lze najít již ve starém Řecku. Jednu z prvních myšlenek vyslovil Anaximandros (6. st. př. n. l). V 5. st. př. n. l. přišel řecký filozof Empedoklés s teorií vzniku života, podle které z prvotního chaosu vznikly jednotlivé části rostlinných i živočišných těl, které se navzájem různě spojovaly a kombinovaly, až vznikly normální organismy. Aristoteles (4. st. př. n. l.) promýšlel podrobnou teorii postupného vývoje světa. Domníval se, že ke vzniku orgánů či organismů je nezbytná finální příčina, která orgánu nebo organismu dává účel.
Vzhledem k význačné církevní moci ve středověku bylo zpochybňování stvoření světa existenčně nebezpečné. Proto se s prvními evolučními teoriemi setkáme až na konci 17. století, kdy církevní vliv již upadá. Evoluci se věnovalo hned několik anglických, francouzských i německých vědců a filozofů. Škola diluvianistů (diluvium = potopa) vznikla na začátku 18. století; její zastánci považovali nalezené zkameněliny za zbytky živočichů, kteří zahynuli při biblické potopě světa. Položili tak základy paleontologie (= nauka o vyhynulých zvířatech a rostlinách). Anglický přírodovědec R. Hooke zastával názor, že se potopa několikrát opakovala. Na začátku 19. století se paleontologie stala jedním z biologických oborů. Založil ji francouzský přírodovědec G. Cuvier, který zastával podobný názor jako Hooke. Cuvier změny fauny a flóry vysvětloval hromadným zánikem při katastrofách světa a novým stvořením jiných forem (teorie kataklysmat – celosvětových katastrof). Tato teorie částečně vyhovovala církvi, neboť vysvětlovala nálezy prehistorických zvířat, jako pozůstatky tvorů, kteří žili před potopou světa, při níž zahynuli.
První ucelenou evoluční teorii poskytnul až lamarckismus, jehož zakladatelem byl francouzský přírodovědec Jean-Baptiste de Lamarck (1744–1829). Tvrdil, že živé pochází z neživého, že složitější organismy vznikají v procesu historického vývoje z jednoduchých. Za hybnou sílu vývoje pokládal změny prostředí, evokující aktivní potřebu organismů měnit se v dokonalejší a složitější formy, a to pomocí vůle a vrozených schopností. Vylepšení, která si tak organismus za svého života vytvořil, předává svým potomkům (například žirafa se natahuje do větví, proto budou mít potomci delší krky, potomci rychle běhající srny budou mít silné nohy, synové kovářů budou svalnatější). Toto pojetí (dědičnost získaných vlastností) později zatlačila do pozadí teorie Darwinova. Definitivně byly Lamarckovy představy o evoluci odmítnuty poté, co se ve 20. století podařilo získat více informací o rozmnožování a dědičnosti organismů. Po jistou dobu byly tyto myšlenky v extrémní zpolitizované formě vzkříšeny v SSSR a jeho satelitech jako tzv. lysenkismus.

 Obr. Darwinův předchůdce Lamarck se domníval, že zvířata mají vnitřní potřebu přizpůsobovat se prostředí, jejich orgány používáním mohutní a takto získané vlastnosti se přenášejí na potomstvo. Klasickým příkladem pro něj byly žirafy natahující původně krátký krk ve snaze dosáhnout na listí v korunách stromů. Na snímku pralesní žirafa okapi a žirafa v savaně, využívající potravní niku nedostupnou menším býložravcům (Foto LP).
Obr. Darwinův předchůdce Lamarck se domníval, že zvířata mají vnitřní potřebu přizpůsobovat se prostředí, jejich orgány používáním mohutní a takto získané vlastnosti se přenášejí na potomstvo. Klasickým příkladem pro něj byly žirafy natahující původně krátký krk ve snaze dosáhnout na listí v korunách stromů. Na snímku pralesní žirafa okapi a žirafa v savaně, využívající potravní niku nedostupnou menším býložravcům (Foto LP).
Období klasického darwinismu
Darwinismus představuje evoluční teorii vytvořenou Darwinem a Wallacem. Stal se první evoluční teorií, která obecně ovládla vědecké myšlení. Charles Robert Darwin (1809–1882) byl zakladatelem evoluční biologie. Evoluční teorii opíral o přírodní a pohlavní výběr. V roce 1831 se vydal na bezmála pětiletou výzkumnou cestu kolem světa na lodi Beagle. Během této plavby, zejména při návštěvě souostroví Galapágy, Darwin shromáždil cenná pozorování a bohatý přírodovědecký materiál. Dalšími zdroji inspirace pro něj byly poznatky zakladatele moderní geologie Ch. Lyella o tom, že procesy v dávné minulosti podléhaly týmž zákonitostem jako děje v současnosti, a ekonoma T. R. Malthuse, který ukázal, že zatímco růstová kapacita populací je neomezená, zdroje jsou naopak limitované a v důsledku toho dochází k soutěži o ně, kdy jen malý podíl jedinců se nakonec zapojí do reprodukce. Zásadním zdrojem poznatků bylo i rozvíjející se plemenářství a šlechtitelství v tehdejší Anglii. Darwin postupně uspořádal svou základní koncepci přirozeného vzniku a vývoje druhů evolucí, jejímž hlavním hybatelem je přírodní výběr. Své názory po 25 letech práce shrnul v díle O vzniku druhů přírodním výběrem v roce 1859. Zakladatel biogeografie Alfred Russel Wallace (1823-1913) svou prakticky totožnou teorii evoluce vypracoval současně s Darwinem a nezávisle na něm. Zatímco Lamarck soudil, že dědičné adaptivní změny jsou přímým důsledkem spolupůsobení vnějších faktorů a vnitřní vůle organismu, Darwin vycházel z předpokladu, že adaptace je důsledkem přírodního výběru (selekce) z náhodných odlišností jedinců v populaci a je tedy zcela nezávislá na vůli či snažení organismů. Mimo tohoto přírodního výběru postuloval Darwin ještě pojetí pohlavního výběru, vycházející z myšlenky, že podobu organismu neurčuje pouze jeho schopnost dobře žít, ale též být atraktivní pro své případné sexuální partnery, neboť kdo nenajde partnera k páření, ten nepředá potomkům své vlastnosti. Některé Darwinovy myšlenky jsou dnes již překonány; Darwin např. neměl k dispozici poznatky moderní genetiky (byť byl současníkem Mendela), zejména nevěděl nic o problematice mutací. V jeho době nebyla ještě známa řada tzv. "chybějících článků" ve fosilním záznamu, stáří Země bylo silně podhodnoceno, stěží bylo možno věrohodně vysvětlit evoluci složitých orgánů jako oko nebo mozek. Darwin svým dílem odstranil člověka z privilegovaného postavení mimo ostatní tvorstvo a přírodu (původ člověka "z opice"), což vedlo k ostrému konfliktu s náboženskými autoritami. Dodnes je Darwin v konzervativních a církevních kruzích nejméně tolerovaným vědcem a stále jsou konány pokusy vytlačit evoluční teorii kreacionismem v moderním hávu, např. tzv. teorií inteligentního designu. Darwinovy myšlenky v Anglii obhajoval a prosazoval zejména T. H. Huxley (vysloužil si tak přezdívku "Darwinův buldok") a v Německu vlivný zoolog Ernst Haeckel (1834-1919).
Základní předpoklady Darwinovy teorie zahrnují:
- teorie existence evoluce druhů - druhy nejsou neměnné, ale vyvíjí se v čase
- teorie společného původu druhů - vývoj z jednoho společného předka
- teorie gradualismu - postupný vývoj hromaděním drobných změn
- teorie přirozeného výběru jako hlavního mechanismu pohánějícího veškeré evoluční změny
Období neodarwinismu
Neodarwinismus je souhrnné označení pro současné hlavní paradigma evoluční teorie vzešlé z Darwinova učení. Představuje snahu syntetizovat hlavní myšlenky darwinismu s poznatky genetiky a dalších disciplín. Hlavním stimulem pro jeho vznik bylo znovuobjevení Mendelových zákonů a rozvoj klasické genetiky v prvních desetiletích 20. st. Někdy je pojem neodarwinismus nahrazen pojmem moderní evoluční syntéza. Tento termín poprvé použil Julian Huxley ve své knize Evolution: The Modern Synthesis (1942). Dalšími významnými osobnostmi v tomto novém směru vnímání evoluce jsou Ernst Mayr, Theodosius Dobzhansky, Ronald A. Fisher a další.
Základní předpoklady neodarwinismu zahrnují:
- hlavním mechanismem vzniku druhů je geografická izolace populací
- akceptuje původní darwinistické mechanismy evoluce ve spojení s Mendelovými zákony a paleontologickými poznatky získanými v průběhu 20. st.
- podstatu variability organismů, která je pro přírodní výběr nezbytná, vysvětluje na genetické bázi
Postneodarwinismus
Toto období je také označováno jako období molekulárně-evolučních teorií. Rozvoj molekulární biologie umožnil studium procesů evoluce na úrovni buňky a vypracování nových teorií vzniku a vývoje života. Zdůrazňuje úlohu genového driftu, náhodných změn genotypu, popř. možnost uplatnění neutrálních mutací na úkor působení přírodního výběru. Jedná se o nejmodernější pojetí evoluce, jehož koncepce se datuje na přelom 60. a 70. let a je spojena zejména s Richardem Dawkinsem (nar. 1941) a jeho teorií tzv. sobeckého genu. Na evoluci pohlíží jako na hru jednotlivých alel (nikoliv jedinců, populací či druhů) soutěžích o své přetrvání prostřednictvím fenotypů, které kvůli tomu vytvářejí. Tento gen je oproti jiným alelám replikován častěji a mívá obvykle pozitivní dopad na fitness svého nositele. Objevily se též tzv. teorie evolučně stabilních strategií, jejichž autory jsou J. M. Smith a G. Price. Kritérium úspěšnosti určité biologické vlastnosti není to, jak zvyšuje či snižuje biologickou zdatnost svého nositele, ale to, zda se jedná o evolučně stabilní strategii ve smyslu teorie her. Ukázali, že z dlouhodobého hlediska má šanci pouze taková strategie, která jakmile jednou v populaci převládne, je schopná zabránit invazi kterékoli strategie minoritní. Další zásadní posun v evoluční biologii přinesla práce paleontologů N. E. Eldredga a S. J. Goulda, kteří ukázali na skutečnost, že evoluce druhů má výrazně nespojitý charakter. Druhy se mění bezprostředně po svém vzniku a pro nesrovnatelně delší období jejich existence je charakteristická evoluční stáze.
Základní mechanismy evoluce
Organismy patří mezi systémy s pamětí, které se tedy v průběhu evoluce vyvíjejí. Z hlediska evoluce má význam populace a nikoli jedinec, jelikož jedinec se během života genotypově nemění, brzy umírá, zatímco populace je genotypově variabilní a trvale pokračuje v čase. Nápadnou vlastností živých soustav je jejich složitost (komplexita). Komplexní systémy vznikají v evoluci působením celé řady různých procesů. Předpokladem evoluce je existence genetické variability v populaci. Významnými zdroji této variability jsou mutace a rekombinace genetické informace. Rozdílný přenos genetické variability do dalších generací probíhá prostřednictvím evolučních mechanismů.
Základní mechanismy evoluce zahrnují:
- výběr neboli selekci (přírodní, pohlavní, rodičovský, umělý)
- genetický drift (posun)
- migraci neboli tok genů
- příbuzenské křížení (inbreeding)
Přírodní a pohlavní výběr
Mechanismus aktivního přírodního výběru je možné představit si tak, že struktury evolučně původnější a pravděpodobně i podstatně jednodušší danou funkci organismu plnily hůře, takže jejich nositelé podlehli v konkurenci s jedinci, u kterých vlivem mutace vznikla struktura komplikovanější a zároveň z funkčního hlediska lepší. Naproti tomu existuje i pasivní přírodní výběr, který nezpůsobuje zvyšování funkčnosti struktury, odstraňuje však následky evolučních změn, které by jinak vedly ke zhoršování její funkčnosti. Přirozená selekce je slepý přírodní proces, zatímco umělý výběr je vyvolán člověkem s určitým záměrem, jehož typickým příkladem je šlechtění.
Mezi příslušníky stejného pohlaví dochází ke kompetici o vhodné sexuální partnery provázené zpravidla intenzivním výběrem – výběrem pohlavním. Působením pohlavního výběru jsou selektováni jedinci s větší sexuální zdatností, kteří dokážou získat sexuálního partnera efektivněji než jejich konkurenti. Toto vede mj. k rozdílné evoluci morfologických znaků u obou pohlaví a pohlavnímu dimorfismu. Např. pestrost a dlouhý "ocas" samců páva jsou výsledkem pohlavního výběru (preference samicemi, ačkoli zvyšují riziko predace) a kryptické zbarvení samic je důsledkem přírodního výběru (snižuje riziko predace).
 Kromě přírodního a pohlavního výběru hrají v evoluci významnou úlohu výběr příbuzenský a rodičovský.Příbuzenský výběr vysvětluje altruistické chování sociálních živočichů, které zvyšuje pravděpodobnost přežití blízce příbuzných jedinců (podrobněji viz u včel v kap. o domestikaci). Rodičovský výběr je prováděný rodiči na svých mláďatech (například u krmivých ptáků se předpokládá, že pestře zbarvené vnitřky zobáků a ústní dutiny vznikly proto, že rodiče přednostně krmili mláďata s nápadnějšími, a tedy barevnějšími vnitřky zobáků). Předpokládá se, že velká část altruistického chování, s nímž se v přírodě setkáváme, vznikla výběrem příbuzenským, a je tedy primárně určena na pomoc blízkým příbuzným altruistického jedince.
Kromě přírodního a pohlavního výběru hrají v evoluci významnou úlohu výběr příbuzenský a rodičovský.Příbuzenský výběr vysvětluje altruistické chování sociálních živočichů, které zvyšuje pravděpodobnost přežití blízce příbuzných jedinců (podrobněji viz u včel v kap. o domestikaci). Rodičovský výběr je prováděný rodiči na svých mláďatech (například u krmivých ptáků se předpokládá, že pestře zbarvené vnitřky zobáků a ústní dutiny vznikly proto, že rodiče přednostně krmili mláďata s nápadnějšími, a tedy barevnějšími vnitřky zobáků). Předpokládá se, že velká část altruistického chování, s nímž se v přírodě setkáváme, vznikla výběrem příbuzenským, a je tedy primárně určena na pomoc blízkým příbuzným altruistického jedince.
Genetický drift
Genetickým driftem rozumíme náhodné posuny ve frekvenci jednotlivých alel v genofondu určité populace. Ukazuje se, že právě genetický drift může za určitých podmínek významně ovlivnit průběh biologické evoluce v některých systémech, a to do takové míry, že zvrátí či silně omezí vliv přírodního výběru. Hlavním projevem tohoto posunu je snižování genetického polymorfismu populace v důsledku upřednostnění některých alel oproti jiným. Ke ztrátě některých alel může vést obdobným způsobem tzv. průchod populace „hrdlem láhve“. Nastává v situacích, kdy z nějakého důvodu významně poklesne početnost populace. Početně zredukovaná populace se reprodukuje a představuje tak základ nové, početnější populace, jejíž genetická struktura však bude odlišná od genetické struktury původní populace. Při průchodu populace hrdlem láhve bude pravděpodobnost ztráty větší pro alely vyskytující se ve velmi nízké četnosti (vzácné alely); naopak alely zastoupené v populaci ve vyšší četnosti mají větší pravděpodobnost projít hrdlem láhve. Pokud vzácná alela projde hrdlem láhve, může se poměrně rychle v nové populaci rozšířit. S tím souvisí též tzv. efekt zakladatele. Uplatňuje se zejména v případech, kdy je určitá nika obsazena malým počtem jedinců, kteří dají základ nové populaci.
Genový tok
Předávání genů mezi populacemi probíhá nejčastěji prostřednictvím migrujících jedinců a je velmi důležitým evolučním faktorem. Schopnost migrovat určuje, jak jsou mezi sebou vzdálená místa, kde se určitý jedinec narodil a kde se narodí jeho potomci. To znamená, že mnohem intenzivnější genový tok má populace borovice, jejíž pyl se šíří větrem na velké vzdálenosti, než populace netopýra, jejíž příslušníci sice za život nalétají tisíce kilometrů, nicméně nakonec odchovají mláďata ve stejné jeskyni, v níž se narodili.
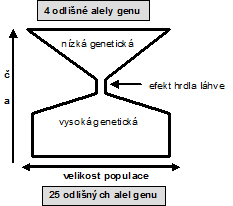
 Obr. Gepard (Acinonyx jubatus) a efekt hrdla láhve. Jako efekt hrdla láhve se označuje stav, který nastane při prudkém poklesu počtu jedinců v populaci. Dochází k příbuzenskému křížení a nevratnému snížení genetické variability. Výzkumy ukázaly, že k něčemu takovému došlo u gepardů asi před 10 tis. lety. Všichni žijící gepardi jsou si vzájemně velmi příbuzní, a to do té míry, že kožní štěp přenesený z jednoho geparda na jiného není odmítnut imunitním systémem. Vzory kreseb gepardů z různých koutů Afriky se od sebe prakticky neliší. Vlivem příbuzenského křížení klesá vitalita mláďat a oplozovací schopnost spermií. Počet spermií v ejakulátu geparda je o 90% nižší než u lva nebo tygra, a 75% z nich je nějakým způsobem abnormální (mají dva nebo více bičíků, žádný bičík, deformovanou hlavičku atd.). Budoucnost gepardů je tedy značně nejistá.
Obr. Gepard (Acinonyx jubatus) a efekt hrdla láhve. Jako efekt hrdla láhve se označuje stav, který nastane při prudkém poklesu počtu jedinců v populaci. Dochází k příbuzenskému křížení a nevratnému snížení genetické variability. Výzkumy ukázaly, že k něčemu takovému došlo u gepardů asi před 10 tis. lety. Všichni žijící gepardi jsou si vzájemně velmi příbuzní, a to do té míry, že kožní štěp přenesený z jednoho geparda na jiného není odmítnut imunitním systémem. Vzory kreseb gepardů z různých koutů Afriky se od sebe prakticky neliší. Vlivem příbuzenského křížení klesá vitalita mláďat a oplozovací schopnost spermií. Počet spermií v ejakulátu geparda je o 90% nižší než u lva nebo tygra, a 75% z nich je nějakým způsobem abnormální (mají dva nebo více bičíků, žádný bičík, deformovanou hlavičku atd.). Budoucnost gepardů je tedy značně nejistá.
Evoluční tah
Evolučním tahem se rozumí evoluční vývoj genetické informace realizovaný na úrovni genoforu. Mezi základní typy evolučních tahů patří: molekulární, mutační a reparační. Molekulární tahy se uplatňují při divergenčních procesech, zejména prostřednictvím změn genových četností v genofondu populací. Na rozdíl od genetického driftu nejsou při molekulárním tahu změny genových četností způsobené náhodně, pokud jde o jejich směr. To znamená, že po rozdělení původní velké populace na více subpopulací budou vlivem molekulárního tahu nakonec ve všech subpopulacích fixovány tytéž alely (na rozdíl od genetického driftu, který vede k fixaci různých alel v jednotlivých subpopulacích) a to bez ohledu na dopad na reprodukční zdatnost jejich nositelů. Molekulárními tahy se patrně v genofondu populací efektivně rozšiřuje tzv. sobecká DNA, tj. zpravidla vysoce repetitivní nekódující DNA v genomu eukaryotických organismů. Pojmem mutační tah se rozumí preferenční vznik určitých typů mutací v některých místech molekuly DNA, resp. RNA a naopak reparační tah je důsledkem reparace genoforu (mechanismem je mutace, která vede k opravě genu).
Mikroevoluce, speciace, makroevoluce
Evoluční procesy bývají klasifikovány podle různých kritérií. Nejčastěji se používá jako kritérium úroveň taxonomické kategorie, podle něhož se rozlišují tři úrovně biologické evoluce: mikroevoluce – týká se evolučních změn uvnitř druhu; základní jednotkou je zde populace; speciace – týká se změn ve vývojových liniích (zejm. divergence) vedoucích ke vzniku nového druhu; makroevoluce – týká se vzniku vývojových linií s více druhy, tedy jinými slovy, vzniku taxonů vyšších než je druh.
Mikroevoluce
Jde o typ biologické evoluce, která se uskutečňuje za pomoci postupného hromadění se drobných změn v krátkých časových úsecích, například změny frekvencí alel v populacích během několika generací. Tyto změny se uskutečňují prostřednictvím mutace, přírodního a umělého výběru, genetického posunu a toku genů. Pozorovatelnými krátkodobými důsledky mikroevoluce jsou například vývoj rezistence bakterií k antibiotikům, odolnost hmyzu k insekticidům nebo hlodavců k rodenticidům. Dalším příkladem je tzv. průmyslový melanismus u píďalky drsnokřídlece březového (Biston betularia). Typická forma je bílá s černými skvrnami, což ji, pokud sedí na bříze, chrání před predátory. Existuje však i melanická forma, která je tmavá a vznikla díky řídce se vyskytující mutaci. Tato nová forma byla poprvé objevena v roce 1848 v Manchesteru. V letech 1848-1898 v souvislosti s průmyslovou revolucí v dané oblasti její frekvence výskytu značně vzrostla a tato forma se stala běžnou. Faktorem přírodního výběru jsou hmyzožraví ptáci, kteří se orientují zrakem na ty jedince, kteří jsou jinak zbarvení, než podklad, na kterém sedí. V lesích, kde je spad popílku značný a kůra stromů tmavá, je procento přežívajících černých forem větší a naopak v lesích nezasažených spadem je větší procento původních forem. Dlouhodobé důsledky mikroevoluce je možné pozorovat u druhů s rozsáhlým areálem, kdy jednotlivé subpopulace podléhají odlišnému přírodnímu výběru vedoucímu ke vzniku poddruhů (subspecie) případně k vytvoření nových samostatných druhů. V populaci existuje v důsledku přírodního výběru selekce proti suboptimálním fenotypům. Tuto selekci je možné vyjádřit jako selekční tlak, který podle jeho směru působení dělíme na:
- stabilizující (normalizující, centripetální) – směřuje vždy k eliminaci krajních fenotypů (a potažmo genotypů) při téměř se neměnícím populačním průměru a tím ke snižování genetické variability populace
- selekce usměrněná (direktivní) - zvýhodňuje některý z extrémních fenotypů, důsledkem je posun populačního průměru ve směru selekce (tj. k extrémním hodnotám) a snížení genetické variability populace
- selekce disruptivní (diverzifikující, centrifugální) - uplatňuje se tehdy, když se během jedné generace část členů určité výchozí populace dostane do prostředí, v němž je selekcí preferován jeden extrémní fenotyp a druhá část členů populace se dostane do prostředí, v němž je selekcí preferován opačný extrémní fenotyp.
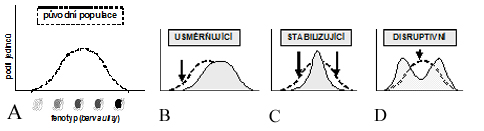 Obr. (A) Vliv usměrňujícího, stabilizujícího a disruptivního výběru určující směr evoluce určitého znaku v populaci. Vysvětleno na příkladu zbarvení ulity plže. (B) Posun celkové skladby populace ve prospěch jedné z extrémních variant barvy ulity. Preference tmavšího zbarvení ulity. Možným vysvětlením je ztmavnutí zemského povrchu v důsledku pokrytí lávou. (C) Eliminace jedinců s extrémním fenotypem – neobvykle světlou nebo naopak tmavou ulitou. Selekce probíhá ve prospěch redukce fenotypové variability. (D) Podporuje varianty obou extrémů proti průměru. Zvyšuje se relativní frekvence výskytu jedinců s velmi světlou nebo naopak s velmi tmavou ulitou. Možným vysvětlením je výskyt ve „skvrnitém“ prostředí (světlý písek hustě pokryt sopečnými kameny).
Jde o proces, v jehož průběhu vzniká ze starého druhu jeden či více druhů nových. Existují dva základní předpoklady pro možnost speciace: existence reprodukčně-izolačních mechanismů (RIM), které jsou schopny přerušit tok genů a existence varibility populací. Reprodukčně-izolační mechanismus je vlastnost, která zabraňuje dvěma druhům, aby se vzájemně křížily (např. geografické nebo časové izolace). Patří sem i např. odlišný zpěv morfologicky velmi podobných příbuzných druhů ptáků (tzv. podvojné druhy, sibling species), rozdílná vokalizace žab, rozdíly v epigamním chování, v načasování reprodukce aj.
Obr. (A) Vliv usměrňujícího, stabilizujícího a disruptivního výběru určující směr evoluce určitého znaku v populaci. Vysvětleno na příkladu zbarvení ulity plže. (B) Posun celkové skladby populace ve prospěch jedné z extrémních variant barvy ulity. Preference tmavšího zbarvení ulity. Možným vysvětlením je ztmavnutí zemského povrchu v důsledku pokrytí lávou. (C) Eliminace jedinců s extrémním fenotypem – neobvykle světlou nebo naopak tmavou ulitou. Selekce probíhá ve prospěch redukce fenotypové variability. (D) Podporuje varianty obou extrémů proti průměru. Zvyšuje se relativní frekvence výskytu jedinců s velmi světlou nebo naopak s velmi tmavou ulitou. Možným vysvětlením je výskyt ve „skvrnitém“ prostředí (světlý písek hustě pokryt sopečnými kameny).
Jde o proces, v jehož průběhu vzniká ze starého druhu jeden či více druhů nových. Existují dva základní předpoklady pro možnost speciace: existence reprodukčně-izolačních mechanismů (RIM), které jsou schopny přerušit tok genů a existence varibility populací. Reprodukčně-izolační mechanismus je vlastnost, která zabraňuje dvěma druhům, aby se vzájemně křížily (např. geografické nebo časové izolace). Patří sem i např. odlišný zpěv morfologicky velmi podobných příbuzných druhů ptáků (tzv. podvojné druhy, sibling species), rozdílná vokalizace žab, rozdíly v epigamním chování, v načasování reprodukce aj.
Speciaci můžeme členit na:
- anagenezi neboli fyletickou speciaci - proces, při kterém dochází ke speciaci tak, že se postupně jeden druh jako celek geneticky mění ve druh jiný (hromadění změn v rámci neštěpících se linií)
- kladogenezi (řec. klados = větev) neboli štěpnou speciaci - proces vzájemného odvětvování (štěpení) vývojových linií, kdy jeden druh mateřský se rozpadne na dva druhy dceřiné, které se dále vyvíjejí samostatně
- syngenezi, při které dochází ke spojování původně samostatných vývojových linií, které se pak mohou dále větvit (Obr.). Existují dva základní mechanismy syngeneze:
- symbiogeneze - vznik nového druhu organismu integrací dvou nepříbuzných organismů, které žijí po určitou dobu v nějaké formě symbiózy, do organismu jediného. Nejznámějším příkladem je vznik eukaryot, k němuž došlo právě postupnou integrací zástupců několika nepříbuzných linií organismů prokaryotických.
- mezidruhová hybridizace - náhodné křížení příslušníků dvou různých druhů. Na rozdíl od symbiogeneze se může uplatnit pouze u blízce příbuzných druhů s pohlavním rozmnožováním. Jako příklad je možné uvést vznik skokana zeleného z rodičovských druhů skokana skřehotavého a skokana krátkonohého. Tato hybridní populace (klepton) je schopna pokračovat v existenci pouze křížením s jedinci rodičovského druhu.
Vznik druhů může dále zahrnovat speciaci alopatrickou a sympatrickou:
-
Alopatrická speciace (řec. allos = jiný, lat. patria = vlast) - nový druh vzniká postupným vývojem mimo přímý kontakt s druhem mateřským. Jestliže například vznikne geograficky izolovaná populace, která se odštěpila od populace mateřské a bude dostatečně dlouho reprodukčně izolovaná, budou se u ní hromadit genetické změny, které povedou k rozrůznění populací.
- alopatrická vikariantní - původně jednotná populace se rozděluje nějakou bariérou (pohoří, řeka) na dvě stejně velké populace a tyto populace se časem geneticky i fenotypově rozrůzní (např. dvojice příbuzných druhů jihoamerických pěvců mravenčíků s výskytem západně a východně od And)
- peripatrická (tzv. efekt zakladatele) - z mateřské populace se oddělí jen malá subpopulace, která dá posléze z důvodu výskytu v odlišných ekologických podmínkách a následné adaptaci vzniknout novému druhu (klasickým příkladem jsou galapážské "Darwinovy pěnkavy", hejnko jejichž předka sem bylo zaváto větry z jihoamerické pevniny)
- Sympatrická speciace - nový druh se formuje na stejném území, na kterém se vyskytuje druh mateřský (např. vznik stovek druhů cichlid ze společného předka obsazováním různých nik v africkém jezeře Malawi, naše druhy rákosníků obsazující různé niky, přesun parazita na jiného hostitele a následná speciace v důsledku přizpůsobení na nový hostitelský organismus).
- Parapatrická speciace – představuje článek mezi alopatrickou a sympatrickou speciací. Oba druhy zaujímají odlišné areály ve vzájemném dotyku; může mezi nimi docházet k toku genů prostřednictvím migrantů. Je typická pro druhy s lineárním rozšířením (např. pobřeží) nebo velkým areálem a tudíž s výrazně rozdílnými životními podmínkami (prostředím). Silný lokální selekční tlak může vést k podstatným změnám v genofondu lokální populace (subpopulace), které nakonec dospějí až ke vzniku nového, samostatného druhu. Takto vzniklo např. několik blízce příbuzných druhů racků s boreálním cirkumpolárním rozšířením (r. stříbřitý, žlutonohý aj.).
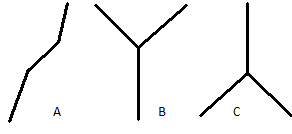 Obr. Speciace: A – anageneze, B – kladogeneze, C – syngeneze.
Obr. Speciace: A – anageneze, B – kladogeneze, C – syngeneze.
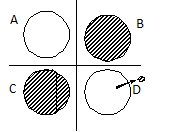 Obr. Klasifikace geografických speciací (Flegr 2005): A – sympatrická, B – parapatrická, C – alopatrická vikariantní, D – peripatrická.
Obr. Klasifikace geografických speciací (Flegr 2005): A – sympatrická, B – parapatrická, C – alopatrická vikariantní, D – peripatrická.
Makroevoluce
Makroevoluce je spojena s většími evolučními změnami probíhajícími v dlouhém časovém úseku. Zahrnuje vznik a evoluci vyšších taxonů než je druh a evoluci novinek (apomorfií), které tyto taxony charakterizují.
Každý živočišný druh dospěl do své podoby dlouhodobou vývojovou cestou. Tomuto vývoji z původních společných předků do dnešních forem říkáme fylogeneze (lat. phylum = kmen, genesis = vznik). Osud vývojových linií může být různorodý. Linie mohou dlouhodobě setrvávat v klidu beze změn (viz dále). Další možností je štěpení linie do velkého počtu druhů během krátkého úseku – adaptivní radiace (lat. radius = paprsek) resp. divergence. Klasickým příkladem jsou opět "Darwinovy pěnkavy". Může docházet i k úpadku a v konečném důsledku extinkci vývojové linie.
Velký význam v evoluci hraje preadaptace. Preadaptace je vznik vlastnosti předtím, než vznikla nebo se uplatnila její současná adaptivní funkce. Podstatné je, že preadaptace vzniká náhodně a v počáteční fázi není vystavena adaptivní selekci. Příkladem velmi významné preadaptace je kutikula primitivních vodních členovců. Hlavní funkcí kutikuly je v tomto případě tvorba vnějšího skeletu pro úpon svalů a tím umožnění efektivnějšího pohybu. Současně však tato kutikula umožňuje po drobných úpravách omezit vysychání organismu – to je vlastnost, která se ovšem ve vodním prostředí neuplatnila a k jejímu využití a zdokonalení došlo následně až v souvislosti s přechodem k suchozemskému způsobu života. Dalším příkladem může být peří u předků ptáků, které původně plnilo různé funkce (sociální signalizace, detoxikace, péče o potomstvo, termoregulace), ale bylo zároveň vhodnou preadaptací k letu.
Velmi účinným mechanismem evoluce je vznik tzv. klíčové inovace, která svým nositelům umožní obsadit novou adaptivní zónu, tj. využít určitý soubor nik, které jim do té doby nebyly přístupné. V případě, že tento soubor nik je velmi rozsáhlý, může dojít k adaptivní radiaci, kdy se příslušná linie rozštěpí na velké množství odlišných druhů, z nichž každý může dát vzniknout novým vývojovým liniím. Velmi často se uplatňuje tzv. efekt pána hory. Jakmile je nějaká ekologická nika obsazena určitým druhem, nemůže být tento druh jednoduše vytěsněn druhem, který sem pronikl později. Výhodou „pána hory“ je jeho početní převaha nad druhem novým a momentálně lepší přizpůsobení lokálním podmínkám.
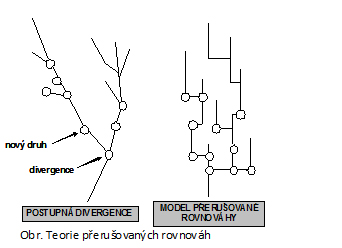
Darwin opakovaně zdůrazňoval, že evoluce je ve své podstatě gradualistická, postupuje střídavě menší nebo větší rychlostí díky postupné kumulaci drobných evolučních změn. Na základě této teorie se předpokládalo, že k většině evolučních změn dochází kdykoliv během existence druhu. Naproti tomu negradualistické – punkcionalistické pojetí evoluce pracuje s teorií přerušovaných rovnováh, která vyzdvihuje skutečnost, že období, ve kterém probíhá speciace, tvoří jen malý zlomek doby existence druhu. Podle této teorie druh velmi rychle vznikl, nabyl svého charakteristického fenotypu a pak se již po dobu své další existence téměř neměnil (Obr.). V tomto případě hovoříme o stazigenezi. Důkazem této dočasné evoluční stagnace jsou např. „živoucí fosílie“ latimérie podivná nebo hatérie novozélandská. Periody intenzivní evoluční aktivity trvající řádově desetitisíce až statisíce let jsou tak střídány stagnací v trvání mnoha milionů let.
Koevoluce
Koevoluce je vzájemná evoluce organismů v těsné evoluční interakci. Koevoluce se řídí vlastními zákonitostmi a v jejím průběhu dochází k řadě jevů, s nimiž se jinak v přírodě nesetkáváme. Koevoluce se velmi často vyskytuje u dvojice dravec-kořist, hostitel-parazit, rostlina-opylovač, u symbiotických druhů, komenzálů, při napodobování různých vzorů (např. za účelem splynutí s okolím - mimikry).
V některých případech je koevoluce dvou druhů těsně provázána a v jejím průběhu vznikají u obou druhů specifická přizpůsobení podmínkám koexistence těchto druhů. Například některé druhy mravenců jsou zcela potravně závislé na medovici vylučované určitým druhem mšic, přičemž tyto mšice jsou závislé na pomoci mravenců při obraně proti nepřátelům nebo na transportu z jedné napadené rostliny na druhou.
Dravec - kořist
V přírodě se setkáváme pouze s těmi druhy, které se dokázaly vyrovnat se skutečností, že se jejich prostředí neustále mění. Musí mít schopnost vlastní evoluce, prostřednictvím které dokáží účelně reagovat na změny prostředí, především na nové evoluční přizpůsobení druhů, s nimiž interagují. Jakmile některý druh nedokáže udržet dostatečné tempo, je eliminován. Příslušný jev popisuje tzv. princip Červené královny, který byl pojmenován podle postavy z knihy Lewise Carrolla „Alenka v kraji za zrcadlem“, jež říká, že „je nutné běžet maximální možnou rychlostí, abychom se alespoň udrželi na místě“. Z tohoto principu například vyplývá, že v přírodě nemohou existovat druhy, které by omezily svou mutační rychlost na nulu. Příkladem tohoto principu jsou tzv. „závody ve zbrojení“ mezi predátorem a jeho kořistí, parazitem a hostitelem, dvěma druhy soupeřícími o využívání určitého zdroje nebo majícími společného predátora. U určitého druhu vzniká evoluční novinka, která zvyšuje jeho zdatnost na úkor příslušníků jiného druhu, a proto se v populaci tohoto druhu postupně rozšíří. Příkladem je vznik silnějších čelistních svalů u dravé ryby, které ji umožní snadněji prolomit schránku plže. Takto vznikne u plže selekční tlak na vytváření pevnějších schránek, který naopak vyvolá selekční tlak na dravou rybu vedoucí ke vzniku vhodné protistrategie, která umožní tyto pevnější schránky rozbíjet. Jiným příkladem koevoluce predátora a jeho kořisti je případ mloka Taricha granulosa a jeho predátora užovky proužkované (Thamnophis sirtalis). Mloci produkují neurotoxin, který se koncentruje v kůži. Užovka si vyvinula rezistenci k tomuto jedu řadou genetických mutací. Vztah mezi těmito dvěma druhy je závodem ve zbrojení vedoucím k extrémní toxicitě mloka a k extrémní odolnosti užovky k tomuto toxinu.
Hostitel - parazit
Zatímco vztah dravec a kořist je výhradně antagonistický, parazit potřebuje živého hostitele jako své životní prostředí a má tedy do určité míry zájmy shodné se zájmy hostitelského organismu. Schopnost proniknout do hostitelského organismu, překonat systémy specifické imunitní obrany hostitele a využívat ho jako životní prostředí a zdroj živin vyžaduje maximální evoluční přizpůsobení parazita hostiteli.
Rychlost evoluce parazita je zpravidla větší než rychlost evoluce jeho hostitele. Zároveň hostitelský organismus na toto přizpůsobení parazita rovněž evolučně reaguje, tudíž evoluce dvojice parazit-hostitel má charakter koevoluce. Stejně jako při evoluci dravec-kořist se i zde setkáváme s velmi intenzivními „závody ve zbrojení“. Evoluce parazita probíhá rychle, parazit produkuje početné potomstvo, z něhož jen část se dožívá reprodukčního věku, uplatňuje se zde tedy přirozený výběr a rychlá evoluce adaptivních znaků. Zároveň další odlišností je mnohonásobně kratší generační doba parazita ve srovnání s hostitelem. Dalším vysvětlením rychlé evoluce parazita je velmi intenzivní selekční tlak ze strany hostitele, kterému je parazit systematicky vystaven.
Pro některé parazitické mikroorganismy jsou například charakteristické tzv. molekulární mimikry, kdy parazit přizpůsobuje strukturu svých makromolekul struktuře příslušných makromolekul hostitelského organismu.
Rostlina - opylovač
Mezi kolibříky a ornitogamními rostlinami (= rostliny opylované ptáky) se vytvořil vztah mutualismu. Pro ornitogamní rostliny je charakteristické nápadné zbarvení květů (odstíny červené, žluté, modré, zelené) a dostatek nektaru s vysokým obsahem cukru, zatímco vůně zpravidla chybí. Na dně květů se shromažďuje nektar, sloužící ptákům jako nápoj. Vztah je výhodný i pro rostlinu, jelikož kolibřík během získávání nektaru zároveň přenáší pyl z květu na květ.
Mimetismus
Vývoj mimetických jevů (též mimikry) je jedním z druhů koevoluce, který je charakteristický napodobováním vzhledu (včetně napodobování chování) příslušníků cizího, často zcela nepříbuzného druhu. Mimetickým vzorem přitom bývají nebezpečné nebo alespoň nejedlé druhy. Rozlišujeme Batesovské a Müllerovské mimikry. Cílem Batesovských mimikry je chránit daného nositele před predátory, kdy jinak neškodný organismus přejímá barevné výstražné znaky jiných, nebezpečných (jedovatých) organismů. Např. pestřenky (dvoukřídlí) napodobují svým vzhledem vosy, včely a čmeláky. V případě müllerovských mimikry jeden nebezpečný či nejedlý druh napodobuje jiný nebezpečný druh proto, aby se jejich společný predátor naučil toto zbarvení lépe rozeznávat. Tím se zvyšuje pravděpodobnost přežití obou lovených druhů (např. neotropičtí motýli rodu Heliconius).
 Obr. Motýli nesytky (Sesiidae) svým vzhledem napodobují nebezpečný blanokřídlý hmyz (Batesovské mimikry). Na Obr. nesytka maliníková (Pennisetia hylaeiformis) (Foto OS).
Obr. Motýli nesytky (Sesiidae) svým vzhledem napodobují nebezpečný blanokřídlý hmyz (Batesovské mimikry). Na Obr. nesytka maliníková (Pennisetia hylaeiformis) (Foto OS).
Člověk jako zdroj evolučních změn
V současnosti se rozhodujícím faktorem v evoluci organismů stal člověk, který svou činností radikálně mění celou biosféru. Činnosti člověka jako možného zdroje evolučních změn můžeme rozdělit do následujících kategorií:
- rychlá změna globálních parametrů biosféry a geosféry, jejichž důsledkem je změna kritérií na výhodné a nevýhodné fenotypy (genotypy)
- změna genetické variability v důsledku zavádění nových genotoxinů (mutagenů)
- přímý zásah do genomu organismů (tzv. geneticky modifikované organismy)
- nebezpečí nukleární energie, která může při masivním uvolnění vyvolat celkové změny v organizovanosti živých soustav a extinkci v globálním měřítku
Zatímco dnešní lidstvo se do značné míry vymklo působení přírodního výběru, člověk již po tisíciletí přetváří umělou selekcí užitkové rostliny a zvířata. Člověk mimoděk mění kritéria pro výhodnost nebo nevýhodnost určitých vlastností a umožňuje tak uplatnění některých druhů (zejména synantropních) na úkor jiných (kulturofobních). Např. již v dávné minulosti kácení lesů prvními zemědělci podpořilo expanzi stepních druhů do naší jinak zalesněné krajiny. Postupem času ovšem novodobé změny ve způsobech hospodaření postihly i tyto druhy, takže mnohé opět téměř vymizely (drop, sysel, koroptev, křepelka). Někteří ptáci se adaptovali ke hnízdění na budovách a prakticky přestali hnízdit původním způsobem ve skalách a stromových dutinách (vlaštovky, jiřičky, rorýsi, některé sovy); mnohé tyto druhy však dnes rapidně mizí v souvislosti s aktuálními změnami (úbytek klasických stájí, zateplování panelových domů, uzavírání vletových otvorů, renovace selských stavení). Kácení deštných pralesů má za následek šíření některých savanových druhů, vysazování monokultur podporuje šíření "škůdců", introdukce nepůvodních druhů je vystavuje novým podmínkám prostředí. Působení člověka je však zatím poměrně krátkodobé a setrvává tak na úrovni mikroevoluce. Naprostá většina živočichů z činnosti člověka rozhodně neprofituje. Dnešní fragmentace ekosystémů by svým izolačním efektem teoreticky mohla napomáhat mikroevoluci a ve velmi dlouhodobém horizontu i speciaci, ve skutečnosti ovšem téměř vždy rychle vede k pravému opaku, tj. mizení druhů, nenávratným extinkcím a globálnímu poklesu a ztrátě biodiverzity. Závěrem je třeba připomenout, že vžitá představa evoluce jako vzestupné, stále se zdokonalující linie resp. "stromu života" na vrcholu s člověkem jakožto pánem tvorstva a vládcem přírody byla snad aktuální za časů E. Haeckela v 19. stol. Naopak, mnohé evoluční linie jsou daleko diverzifikovanější a ekologicky významnější než primáti. Moderní názor na evoluci je takový, že nemá účel a nesměřuje k žádnému určitému cíli. Není na místě ani dělení živočichů na "nižší" a "vyšší". Živočichové resp. jejich vlastnosti jsou buď primitivní (tj. původnější), nebo více odvozené (tj. anageneticky bližší společnému předku nebo od něj vzdálenější); primitivní přitom není totéž co bazální (tj. kladogeneticky bližší společnému předku). Všechny dnes žijící druhy jsou od svého společného předka stejně vzdálené.