ROZMNOŽOVÁNÍ ŽIVOČICHŮ
Nepohlavní rozmnožování
Jinak také asexuální, vegetativní reprodukce. Vznik nového jedince z jedné nebo většího počtu somatických buněk mateřského jedince. Základem je mitotické dělení, genetická informace se přenáší nezměněná, všichni potomci jsou tedy stejní jako generace předchozí. Vyskytuje u skupin bezobratlých s jednodušší tělesnou stavbou a výraznou schopností reparační regenerace (houbovci, žahavci, ploštěnci, kroužkovci, mechovci, chapadlovky, ostnokožci, pláštěnci). Hlavními způsoby nepohlavního rozmnožování jsou dělení a pučení.
Dělení (fisiparie) bývá nejčastěji podvojné (binární). Dceřiní jedinci vznikají rozdělením mateřského jedince, který fakticky zaniká.
U živočichů vzniká nový jedinec až na vzácné výjimky z celého komplexu buněk mateřského organismu. Binární dělení je vždy spojeno s regenerací chybějících orgánů. Při tzv. paratomii dochází k regeneraci ještě před rozdělením jedince.
Video. Nepohlavní rozmnožování naidky chobotnaté (Stylaria lacustris). Díky značné schopnosti regenerace je u kroužkovců časté nepohlavní rozmnožování. Na videu je možné pozorovat jedince, u kterého probíhá binární dělení typu paratomie, kdy dochází k regeneraci chybějících orgánů ještě před rozdělením jedince. Uprostřed naidky lze pozorovat zaškrcení a chobotek nově vznikajícího jedince.
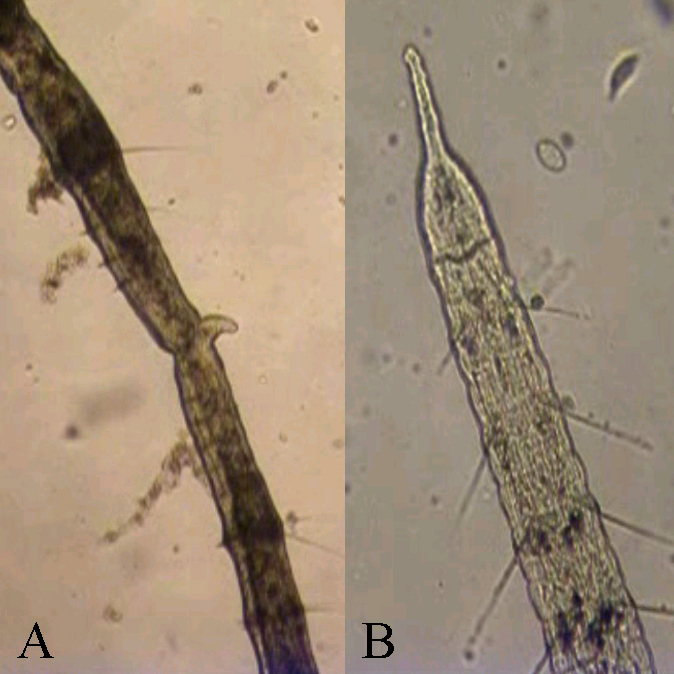
Obr. Detail z předchozího videa zachycující paratomii naidky chobotnaté (Stylaria lacustris ). Uprostřed lze pozorovat zaškrcení a chobotek nově vznikajícího jedince (A). Na druhém snímku je detail hlavy původního jedince (B) (Foto OS).
Formou paratomie, považovanou za přechod mezi dělením a pučením, je strobilace (medúzovci).
Video. Strobilace polypů talířovky ušaté (Aurelia aurita). Příčným zaškrcováním těla přisedlého polypa se nepohlavně tvoří malé medúzky, které volně plavou, rostou a pohlavně dospívají. Převzato z http://www.arkive.org z odkazu na druh Aurelia aurita (funkční adresa 31. 1. 2012).
Střídání pohlavních a nepohlavních generací se obecně nazývá rodozměna (metageneze). Za nedokonalou metagenezi lze považovat epitokii tasemnic nebo mnohoštětinatce palola zeleného ( Eunice viridis), kde nepohlavní generací je atokní část (u tasemnic hlavička, scolex) a neúplnou pohlavní generací epitokní část – články, které nelze považovat za samostatné jedince. U palola od původního nepohlavního jedince (atokní části), žijícího na mořském dně, postupně odděluje řetězec článků (epitokní část) s pohlavními orgány, který po oddělení volně plave u hladiny.
Pokud dceřiní jedinci obdrží jen příslušný podíl tělních orgánů a chybějící části se regenerují až po rozdělení, jde o tzv. architomii. U žížalic (rod Lumbriculus) je znám samovolný rozpad, tzv. metamerická disociace, způsobený prudkými nekoordinovanými pohyby tělní svaloviny, které roztrhají jedince na několik kousků, z nichž každý regeneruje v nového jedince.
K dělení může docházet již v časném embryonálním vývoji, kdy se rýhující vajíčko rozdělí na více samostatných blastomer nebo jejich skupin. Tato tzv. polyembryonie je příležitostná u některých žahavců, máloštětinatců, mechovců, ježovek, ale i u členovců a obratlovců, hlavně u živorodých a parazitických druhů, kde mají zárodky dostatek výživy. U savců je polyembryonie běžná u pásovců (Dasypodidae), u nichž z jednoho vajíčka vzniká rozdělením po stadiu gastruly druhově charakteristický počet 4 až 12 zárodků. U člověka může dojít k fakultativní polyembryonii, a sice při vzniku jednovaječných dvojčat.
Pučení (gemiparie). Při vnějším pučení (gemaci) se na povrchu těla mateřského jedince objevuje pupen, který postupně dorůstá v dceřiného jedince. Ten se časem oddělí a vyvíjí se samostatně, nebo častěji zůstává připojen a tak se postupně vytváří různě rozsáhlé přechodné či trvalé kolonie. Nejčastěji probíhá pučení u přisedlých organismů: u prvoků zejména u nálevníků (např. rournatky – Suctorida); u živočichů především u houbovců, žahavců (polypovců – Hydrozoa, korálnatců – Anthozoa – korálové útesy), mechovců. Obdobně může dojít k pučení vnitřnímu, kdy shluky buněk a později dceřiný jedinec vznikají uvnitř těla jedince mateřského. Gemulací se vyvíjí tzv. klidové pupeny (gemule houbovců, statoblasty mechovců), které slouží k množení a současně k přežívání těchto živočichů v období nepříznivých podmínek. Polyembryonické pučení, tedy pučení v larválním stadiu, známe např. u tasemnic rodu Echinococcus, u kterých se z vaku, vzniklého z larvy onkosféry, vyvíjí stovky menších váčků a v každém z nich pučí dovnitř množství hlaviček, dávajících vznik až tisícům nových tasemnic.
Jinak také sexuální, generativní reprodukce. Nový jedinec se vyvíjí z jediné buňky (zygoty), která vzniká obvykle splynutím samčí a samičí pohlavní buňky (gamety). U primitivních živočichů se vyvíjí pohlavní buňky z buněk somatických. U většiny živočichů však vznikají haploidní gamety složitým procesem (spermatogenezí či oogenezí), jehož základem je redukční meiotické dělení. Pohlavní proces, tedy vývoj, diferenciace a splývání gamet, zabezpečuje rekombinaci genotypu příslušného druhu a tím zvyšuje jeho schopnost přizpůsobovat se změnám životního prostředí. V měnícím se prostředí je pohlavní rozmnožování mnohem výhodnější něž rozmnožování nepohlavní, přestože dochází ke ztrátám při nadprodukci gamet a ke vzniku potomstva jsou nutní dva jedinci místo jednoho. Naopak v konstantním prostředí je nepohlavní množení dobře adaptovaného druhu účinnější než pohlavní. U mnoha druhů se proto vyskytuje kombinace obou způsobů, v závislosti na životních podmínkách (v příznivých, stálých podmínkách se množí nepohlavně; v nepříznivých, proměnlivých podmínkách pohlavně). Toto střídání nepohlavních a pohlavních generací se také často vyskytuje u parazitických živočichů v souvislosti se střídáním hostitele.
U živočichů se v pohlavních orgánech, gonádách, vytváří specializované pohlavní buňky, jde o tzv. oogamii. U gonochoristů jsou samčí gonády (testes) v jiném jedinci než gonády samičí (ovaria). U hermafroditů jsou gonády obojího typu neseny jedním jedincem. Oba typy nalézáme u poměrně blízce příbuzných skupin nejrůznějších bezobratlých (např. nezmar Hydra vulgaris je hermafrodit, Hydra attenuata gonochorista), vzácně i u obratlovců (některé druhy ryb). V případě hermafroditismu mají význam mechanismy zabraňující oplození vajíček vlastními spermiemi, tzv. autofekondaci (vzácně k ní dochází u některých motolic, tasemnic či hlístic). Při simultánním hermafroditismu, kdy gamety dozrávají současně, jsou to oddělené vývody gonád, případně autosterilita, kdy látky vylučované gametami nedovolují kontakt a splynutí gamet téhož jedince (máloštětinatci, pijavice, měkkýši, pláštěnci). Druhým mechanismem je tzv. sukcesní hermafroditismus, při kterém je v určitém období života produkován jen jeden typ gamet, zpravidla mladší jedinci produkují spermie a teprve starší jedinci produkují vajíčka (proterandrie – měkkýši, někteří korýši). Opačný, vzácnější případ se nazývá proterogynie (měkkýši, pláštěnci). Obecně se hermafroditismus vyskytuje u živočichů s malou hustotou populace, jako je tomu v prostorách mořských hlubin a u živočichů s přisedlým způsobem života v dospělosti. Hojný je u parazitů, tedy všude tam, kde je malá pravděpodobnost, že se setkají dva jedinci různého pohlaví.
U gonochoristů se samci a samice liší svými gonádami, tj. primárními pohlavními znaky. Zpravidla se více či méně liší také řadou anatomických a morfologických sekundárních pohlavních znaků, jejichž přítomnost je závislá na znaku primárním. Tyto znaky mohou sloužit přímo rozmnožovací funkci (kopulační orgány, kladélka), vzájemnému vyhledávání, popřípadě stimulaci druhého pohlaví (chemické, optické, akustické signály, svatební tance) nebo péči o potomstvo (břišní vak vačnatců, mléčná žláza). Výrazně odlišné jsou také reprodukční strategie samce a samice (zejména u obratlovců). Samice má oproti samci menší počet gamet a tím i omezené množství potomků. Strategií samice tedy je maximalizovat úspěch v rozmnožování (reprodukční zdatnost, fitness) zvýšením pravděpodobnosti přežití svých potomků. To lze realizovat např. donošením zárodku ve vlastním těle, stavbou hnízda, péčí o mláďata, obranou před nepřáteli, ale také tím, že pro své potomky vybírá co nejkvalitnější otce. Naproti tomu samec produkuje velké množství gamet a jeho strategií je maximalizovat počet svých potomků pářením s co největším počtem samic. Odlišnost jedinců různého pohlaví téhož druhu, pohlavní dimorfismus, se v různé míře vyskytuje u všech skupin živočichů. Výjimkou jsou živočichové přisedlí, málo pohybliví, např. žahavci nebo ostnokožci, jejichž vajíčka jsou oplozována volně ve vodě a jedinci opačného pohlaví se nevyhledávají. Extrémní rozdíly ve velikosti a stavbě těla nalézáme u tzv. komplementárních samečků, kteří jsou miniaturní, mají velmi zjednodušenou stavbu, jsou specializování jen k pohlavní funkci a žijí trvale přichyceni na několikanásobně větší samici či jako komenzálové přímo v jejích pohlavních vývodech. Takový pár se podobá simultánnímu hermafroditu s autofekondací, avšak obě pohlaví pocházejí zpravidla z různých rodičů, takže variabilita a adaptace nejsou na rozdíl od autofekondace redukovány. Komplementární samečkové jsou známi zejména u některých hlubokomořských živočichů (hlavonožců, korýšů, rypohlavce Bonellia viridis, ryb čeledi Ceratiidae z řádu ďasů), u živočichů parazitických (některých klanonožců, stejnonožců, hlístic) i jiných (např. svijonožců, některých pavouků, roztočů).
Druhotné pohlavní znaky mohou být trvalé, ale též periodické, sezónní, zvýrazňující se pouze v období rozmnožování, v době říje (svatební šat, ale i zvláštní chování jako souboje samců, mateřský instinkt aj.). V říji ( oestrus, estrus) dochází k dozrávání a uvolňování gamet. U většiny živočichů probíhá opakovaně, pouze u hmyzu a některých ryb jen jednou za život. Říje většinou souvisí s ovulací a probíhá jednou ročně u monoestrických živočichů (tetřev, orel, netopýři, liška, vlk, srnec, jelen). Také spermie samců dozrávají jen v tomto období. Vícekrát ročně říje probíhá u polyestrických živočichů (většina pěvců, hlodavci, zajíc, domestikovaná zvířata), u kterých bývá spermatogeneze téměř plynulá a samci jsou schopni oplodnit samice kdykoliv.

Obr. Pohlavní rozmnožování je v měnícím se prostředí výhodnější než rozmnožování nepohlavní. U amniot dochází k vnitřnímu oplození (fertilizaci) po páření (kopulace, coitus) v době říje (estrus, oestrus). U většiny fen se říje dostavuje 2krát ročně (diestrické zvíře, nezaměňovat s pojmem diestrus, což je fáze pohlavního cyklu) (Foto IL, Peru).
Cykličnost je ovlivněna ročním obdobím, sezónními změnami (teplota, světlo, výživa) nebo je regulována vnitřním cyklem pohlavních orgánů (pohlavní hormony), jenž ovšem bývá rovněž pod vlivem výše uvedených vnějších faktorů. Regulační mechanismy řídí cykly tak, aby pohlavní buňky dozrávaly současně a aby potomstvo mělo nejvhodnější podmínky pro vývoj, to znamená aby se mláďata líhla nebo rodila v době dostatku potravy (v našich podmínkách na jaře). Proto může být u savců doba mezi pářením a porodem prodloužena. Jedním ze způsobů je tzv. utajená březost (latentní gravidita, oddálená implantace), kdy se vajíčko začne po oplození vyvíjet, ale v určitém stadiu, zpravidla na konci rýhování ve stadiu blastocysty, se vývoj na různě dlouhou dobu zastaví (některé kunovité a medvědovité šelmy, pásovci, srna aj.). V druhém případě, při tzv. utajeném oplození, probíhá říje určitou dobu před ovulací a spermie jsou po kopulaci uchovávány v pohlavních cestách samice až do ovulace, kdy dojde k oplození (u skotu nastává ovulace asi 12 hodin po říji a páření, u netopýrů mírného a subtropického pásma až po půl roce – kopulace v září, říjnu a ovulace v dubnu). U některých savců s poměrně dlouhou dobou říje je známa tzv. indukovaná ovulace (provokovaná říje), kdy stimulem pro ovulaci je vlastní akt páření (králík, zajíc, myš, drobné šelmy). Stimulem může být také ztráta mláďat (případy infanticidy u lvů nebo hulmanů, kdy nový vůdce smečky či tlupy vyvolá zabitím mláďat původního samce u samic říji a spářením s nimi si tak zajistí otcovství). Další úlohou regulačních mechanismů je zabránit dalšímu uvolňování vajíček z vaječníku, jakmile jich je vyprodukováno příslušné množství. Nejvyšší bývá u parazitů – např. u tasemnice Taenia solium asi 42 milionů; nejnižší u živočichů pečujících o vajíčka a mláďata – u tzv. uniparních savců obvykle ovuluje jediné vajíčko (člověk, lidoopi, lichokopytníci, tur, slon), u multiparních více vajíček (ovce, koza, prase, šelmy, hlodavci). Vyloučit další ovulace je významné zejména při vývoji uvnitř těla matky, jakmile dojde k oplození vajíčka. Přesto však přítomnost vyvíjejícího se zárodku u některých živočichů další ovulace nevylučuje. U živorodého žraloka rodu Lamna dochází k pravidelným ovulacím a přítomný zárodek vajíčka polyká (tzv. trofická vajíčka), u halančíkovitých ryb (čeleď Aplocheilidae) může být v pohlavním traktu samice i osm různě starých skupin zárodků (tzv. superfetace). U některých klokanů může mít samice mládě ve vaku, nidovaný zárodek v děloze, oplozená vajíčka čekající na nidaci ve vejcovodech a ve vaječníku vajíčka připravená k ovulaci.
K oplození, tedy splývání gamet, dochází u živočichů ve vodním prostředí (oplození vnější), nebo uvnitř těla samice (oplození vnitřní). U živočichů s vnějším oplozením jsou pohlavní buňky vypouštěny buď necíleně do vody, pravděpodobnost setkání gamet je zvýšena synchronizovanou produkcí obrovského množství těchto buněk (žahavci, ostnokožci), nebo jsou častěji vypouštěny na určitých místech a v určitém čase do blízkosti buněk opačného pohlaví při těsném kontaktu obou jedinců (tření ryb, amplexus žab). Při vnitřním oplození mohou spermie aktivně pronikat z vody do pohlavních orgánů samice, která vajíčka do vody nevypouští (některé ploštěnky, ostnokožci, pláštěnci aj.). Častěji dochází k páření, spermie jsou zaváděny do pohlavních ústrojí samice přímo pomocí pářicích orgánů (penis u savců, některých ptáků, plazů, ryb, u hmyzu aj.; modifikované přední končetiny nebo hlavové přívěsky u pavouků, roztočů či buchanek; hektokotylové rameno u chobotnic). V některých případech samec pomocí penisu, opatřeného bodci, provrtává pokožku samice a spermie aktivně pronikají z parenchymu k ovariu (některé ploštěnky, vířníci, vrtejši). U některých mnohoštětinatců (např. rod Platynereis) vypouští samec sperma mezi čelisti samice a spermie pronikají k vajíčku přes jícen a tělní dutinu. U mnohých živočichů se vyskytují tzv. spermatofory, rosolovité schránky obsahující spermie. Ty samec klade do blízkosti samice, která je přijímá do vývodních cest svých pohlavních orgánů (plži, roztoči, hmyz, čolci a mloci) nebo je samec vkládá do těla samice (hlavonožci).
Případy, kdy se jedinec vyvíjí bez oplození z neoplozeného vajíčka, se označují jako apomixe. Vyskytuje se u některých živočichů, kteří se vzhledem k složité tělesné organizaci nemohou již rozmnožovat nepohlavně. Nejčastější je tzv. partenogeneze, vývoj vajíčka zcela bez účasti spermie (vířníci, kroužkovci, měkkýši, korýši, stonožky, pavoukovci, hmyz, některé druhy ryb a ještěrů). Jde o výhodnou adaptaci sloužící k rychlému namnožení jedinců. Může být jediným způsobem rozmnožování (někteří vířníci, korýši, ještěrka Lacerta saxicola armeniaca), příležitostná nebo se může cyklicky střídat s bisexuálním rozmnožováním – heterogonie (perloočky, mšice, žlabatky aj.). Jiným typem apomixe je gynogeneze (merospermie), při níž je vývoj vajíčka aktivován spermií, jež poskytuje dělicí aparát, který vajíčko ztratilo při redukčním dělení. Ke splynutí jader (karyogamii) však nedojde, jádro spermie zaniká a dalšího vývoje se neúčastní (hlístice, ploštěnky). K aktivaci vajíčka může sloužit dokonce výjimečně i spermie zcela jiného druhu (někteří brouci, vzácně ještěrky a ryby). Např. karas stříbřitý se v původní oblasti výskytu na Dálném východě rozmnožuje bisexuálně, u nás jsou však byly donedávna známy pouze samice, jejichž vajíčka jsou aktivována spermiemi jiných kaprovitých ryb. Další modifikací pohlavního rozmnožování umožňující rychlé střídání generací a prudký růst početnosti je neotenie, schopnost rozmnožování larválních nebo juvenilních stadií (hmyz, obojživelníci - axolotli, macarát jeskynní). Kombinací neotenie a partenogeneze je tzv. pedogeneze (motolice, dvoukřídlí – bejlomorky, pakomáři). Z oplozených vajíček bejlomorek se vyvíjí larvy, které ještě před metamorfózou dávají živorodostí vznik dalším larvám. Tyto se vyvíjejí partenogeneticky a nezřídka se živí mateřskou larvou, přičemž se v jejich těle již vyvíjí další embrya. Při změně životních podmínek se vzniklé larvy zakuklí a dají vznik dospělým samcům a samicím.
U většiny živočichů se jedinci opačného pohlaví aktivně vyhledávají. K vábení a vybírání partnerů slouží celá řada rozmanitých strategií, založených na akustických (zpěv ptáků, zvuky hmyzu, savců), optických (zbarvení, zvláštní typy chování) a chemických (feromony) signálech. Sdružování může mít různý charakter. Většinou se jedinci sdružují jen na dobu páření. U obratlovců se může vytvářet více či méně trvalé soužití jedné samice s jedním samcem (monogamie), přetrvávající i po období rozmnožování v souvislosti se společnou péčí o potomky (liška, většina ptáků – sezónní monogamie). Trvalá monogamie je však poměrně vzácná (vrubozobí, albatrosi, giboni). Naopak častá je u živočichů promiskuita, páření jedinců obou pohlaví s více jedinci druhého pohlaví (živočichové žijící ve větších skupinách). Páření s jinými jedinci je známo i u monogamních druhů jako tzv. extrapárové kopulace, pomocí kterých mohou jedinci zvyšovat svou fitness. Při polygamii se jedinec jednoho pohlaví páří s větším množstvím jedinců druhého pohlaví. Častější je polygynie, kdy se samec páří s více samicemi. Samec si buď vytváří harém, který brání proti dalším samcům (nandu, tinamy, jelen, rypouš sloní), nebo láká samice na určité místo (tokaniště, lek, říjiště), na kterém se samicím předvádí (tetřívci, rajky, lemčíci, daněk). Vzácnější je polyandrie, kdy se samice v období říje páří s větším počtem samců, kteří jí pak mohou vypomáhat s péčí o potomstvo (někteří bahňáci, kukačky). Většinou se v době rozmnožování setkávají dva jedinci. Některé druhy se však shlukují do větších skupin (svatební roje hmyzu) a mohou podnikat i dlouhé reprodukční migrace (anadromní či katadromní druhy ryb, obojživelníci, mořské želvy).