Základní rysy stavby těla obratlovců
Základní rysy stavby těla obratlovců
Somatický skelet (vnitřní kostra) a kosterní svalovina. U většiny dospělých obratlovců (kromě mihulí) je chorda dorsalis silně redukovaná až zcela zaniká (poprvé u ptáků). Chrupavčitá struna hřbetní je postupně nahrazena páteří tvořenou obratli. Vnitřní kostra obratlovců se dá rozčlenit na kostru hlavy, osní kostru a kostru končetin.
Kostra hlavy - lebka (cranium) vytváří schránku sloužící především jako ochrana mozku. Cranium je tvořeno dvěma částmi, v jedné je uložen mozek a nazývá se neurocranium (část mozková), druhou část tvoří viscerocranium neboli splanchnocranium (část obličejová).
Základem osní (axiální) kostry je páteř – columna vertebralis, která je tvořena obratli – vertebrae. Během evoluce docházelo ke vzniku osifikačních center v pravidelných úsecích kolem chordy dorsalis. Později se z těchto center začaly utvářet osifikované prstence, které postupně čím dál více zaškrcovaly strunu hřbetní. Nakonec osifikovaná tkáň obratlových center chordu zcela přerušila na řadu izolovaných krátkých částí, které zůstaly uložené v meziobratlových prostorech (např. nucleus pulposus u savců). Pozůstatky chordy v meziobratlových prostorech formovaly čela sousedních obratlových center do konkávního tvaru. Na základě rozdílného tvarování čel obratlových center vzniklo několik typů obratlových těl. Jestliže bylo obratlové centrum formováno na svém předním (kraniálním) i zadním (kaudálním) konci, označuje se takové bikonkávní tělo obratle jako amficélní. Pokud je tělo obratle na své kaudální straně konkávní a na kraniální konvexní, nazývá se opistocélní. Opačné utváření (na kaudální straně konvexní a na kraniální konkávní) těla obratle je označováno jako procélní. Platycélní tělo obratle je na obou koncích rovné a heterocélní je s přední i zadní plochou částečně konvexní a částečně konkávní.

Obr. Amficélní obratel ryb (Foto OS).
Kost je zřejmě původnější opornou tkání dospělců než chrupavka, která se uplatňuje zejména u zárodků a mláďat. Ve fylogenezi se patrně zakládala úplněji osifikovaná kostra a teprve později se druhotně zvyšoval podíl chrupavky na úkor kosti (mihule, paryby). K hrudní části páteře jsou kloubně připojena žebra – costae. U většiny suchozemských obratlovců je převážná většina žeber ve ventrální části spojena pomocí hrudní kosti – sternum. Vytvořením tohoto kloubního spojení vzniká hrudní koš – thorax, který uzavírá a chrání orgány dutiny hrudní.
Kostra párových končetin se zakládá z trupového mezodermu v podobě končetinových pupenů. U vodních živočichů s ploutvemi se tyto končetinové základy tvoří v rozsáhlejších úsecích odpovídajících místu budoucích ploutví. U suchozemských tetrapodů se končetinové pupeny embryonálně zakládají podél celé délky těla v podobě horizontální lišty (Wolffova lišta). V pozdějších fázích vývoje tato lišta v místech budoucích párových končetin expanduje a v ostatních částech zaniká. Kostra končetin je tvořena pletencem – cingulum a kostrou volné končetiny. Pletenec hrudní končetiny tvoří lopatka – scapula, os coracoideum (procoracoid, krkavčí kost, zobcovitá kost), klíční kost - clavicula. Coracoid je u savců plně vyvinut jen u ptakořitných, u ostatních savců přirůstá její rudiment k lopatce jako processus coracoideus. U živočichů s vyvinutou klíční kostí (u savců pouze u primátů a býv. hmyzožravců) dochází k spojení hrudní končetiny s axiální kostrou v oblasti sterna. Clavicula tvoří oporu hrudní končetině při namáhavé abdukci, tedy při uchopování, hrabání a létání. Pletenec pánevní končetiny – cingulum membri pelvini tvoří kost kyčelní – os ilium, kost sedací – os ischii a kost stydká – os pubis. Pánevní pletenec je na axiální skelet pevně připojen v oblasti křížové kosti – sacrum. Kostru volné hrudní končetiny tvoří kost pažní – humerus, kost vřetenní – radius a loketní – ulna, dále skupina kůstek zápěstních – ossa carpalia, záprstních – ossa metacarpalia a články prstů – ossa digitorum. Analogicky je tvořena pánevní končetina kostí stehenní – femur, kostí holenní – tibia, lýtkovou – fibula a skupinami kůstek zánártních – ossa tarsalia, nártních – ossa metatarsalia a články prstů. Končetiny vodních obratlovců mají tvar ploutví s rozdílným kosterním základem. U suchozemských živočichů se embryonálně zakládají pětiprsté končetiny. Během evoluce, kdy docházelo k přizpůsobování se končetin k různým druhům pohybu, došlo u některých obratlovců k redukci počtu prstů.
Svaly párových končetin (včetně ploutví) mají svůj původ v myotomech somitů mezodermálního původu a vznikají v končetinovém základu současně se skeletem. U vodních obratlovců jsou svaly z větší části uspořádány jednoduše v podobě segmentů – myomery, které jsou od sebe odděleny septy – myosepta (myocommata). V těchto přepážkách u některých ryb částečně osifikuje vazivo a vznikají takzvané kostice. U plazů zůstávají částečně segmentovány pouze dorzální svaly trupu. U pokročilejších suchozemských obratlovců se svalová segmentace vytrácí a lze ji rekonstruovat jen podle uspořádání inervace. U těchto živočichů došlo k vytvoření složitých svalových skupin, které zajišťují pohyb jednotlivých částí těla.
Nervová soustava. Významným znakem obratlovců je nápadný rozvoj nervové soustavy, která se člení na centrální nervovou soustavu (CNS) a periferní nervovou soustavu. Z embryonálního neuroektodermu vzniká u obratlovců podstatná část centrální nervové soustavy, jejíž hlavová část zbytňuje v důsledku kumulace neurální hmoty ve směru převládajícího pohybu do podoby mozku – encephalon, syn. cerebrum. U dospělých obratlovců je mozek členěn na pět oddílů. V přední části je telencephalon - koncový mozek, za ním diencephalon – mezimozek, mesencephalon – střední mozek, metencephalon, syn. cerebellum – mozeček a na přechodu mozku a míchy je myelencephalon, syn. medulla oblongata – prodloužená mícha. Z mozku vystupuje 8–12 párů mozkových nervů. Na rozdíl od míchy je v mozku uložena šedá hmota, tvořená z těl neuronů, na povrchu, zatímco bílá hmota tvořená nervovými vlákny je uvnitř. Mícha - medulla spinalis prochází páteřním kanálem a z každého jejího segmentu vystupují dva páry míšních nervů (kořeny míšní). Nervová vlákna tvořící dorzální kořeny míšní mají senzitivní funkci (dostředivá dráha). Ventrální kořeny míšní obsahují vlákna s funkcí motorickou (odstředivá dráha). Vegetativní (autonomní) nervstvo je tvořeno dvěma víceméně antagonisticky (protichůdně) působícími systémy - sympatikem a parasympatikem. Míšní a mozkové nervy, stejně jako vegetativní nervový systém, tvoří společně periferní nervstvo.

Obr. Evoluce mozku obratlovců. Žlutě – čichový lalok, červeně – optický lalok, šedě – cerebrum, modře – mozeček. Je patrné zvětšování cerebra, u savců i jeho gyrifikace. Mozeček je nejvíce vyvinut u skupin, kde hraje významnou roli rovnováha a jemná koordinace pohybů (ptáci, ryby, savci). (JK podle Hickmana et al., 2004).
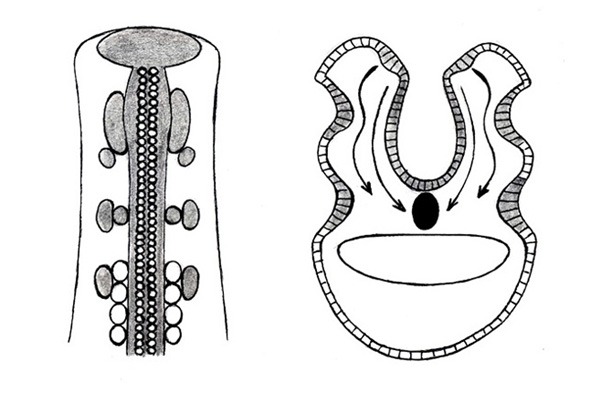
Obr. Plakody a neurální lišta v hlavové části embrya obratlovce (vlevo pohled shora, vpravo příčný průřez). Obě struktury jsou ektodermálního původu; z plakod vznikají smyslové neurony mozku a smyslové orgány a z neurální lišty se odvozuje na 40 tkání a orgánů (lebka, zuby, žaberní aparát, dermální skelet, periferní nervový systém, některé hladké svaly a žlázy aj.). (JK podle Zrzavého, 2006).
Jednotlivé funkce probíhající v organismu obratlovců jsou koordinovány nejen nervovou soustavou, kde vzruchy probíhají usměrněně v síti nervových vláken, ale také prostřednictvím chemických látek – hormonů. Tyto jsou tvořeny v žlázách s vnitřní sekrecí (endokrinní systém) a tekutinami oběhového systému jsou dále šířeny po těle. Chemický přenos vzruchů je patrně evolučně starší, protože se s ním setkáváme již u hub, kde morfologicky diferencovaná nervová soustava ještě chybí.
Smyslové orgány. Mohutný rozvoj smyslových orgánů u obratlovců pravděpodobně souvisí se zvýšeným pohybem těchto živočichů a s tím spojenou nutností orientace a vnímání vnějších podnětů. Smyslové buňky a smyslové orgány, které registrují podněty z vnějšího prostředí, jsou v důsledku toho většinou derivátem vnějšího zárodečného listu (ektodermu). Výjimkou jsou proprioreceptory, které registrují podněty od vnitřních orgánů. Už u evolučně nižších živočichů existuje řada jednoduchých receptorů a některé z nich se dochovaly i u obratlovců. Jsou to například jednoduché georeceptory – smyslové orgány jimiž živočich vnímá svoji polohu. Georeceptory jsou u obratlovců vyvinuty jako pár orgánů na každé straně těla v podobě blanitého labyrintu, umístěného ve vnitřním uchu. Blanitý labyrint je u čelistnatců tvořen dvěma váčky zvanými utriculus a sacculus. Z báze sacculu vybíhá třetí váček zvaný lagena, která je základem hlemýždě – cochlea u savců. V každém váčku je okrsek obrveného smyslového epitelu a váčky jsou vyplněny endolymfou. V této tekutině se vznáší různý počet statolitů (otolitů) v podobě zrníček uhličitanu vápenatého. U paprskoploutvých ryb vyplňuje otolit prakticky celou dutinu sacculu a jeho tvar je charakteristický nejen pro rody, ale i pro druhy, takže se často používá v systematice ryb. Vnímání pohybu a jeho změn (směr a zrychlení) zajišťují polokruhové kanálky – ductus semicirculares. Tyto kanálky vybíhají dorzálně z utriculu, s nímž jsou na obou koncích spojeny a u čelistnatců jsou tři (u mihulí pouze dva). Kanálky leží v navzájem kolmých rovinách a každý je na jednom konci rozšířen v tzv. ampulu. V ampulách je na stěně hřeben smyslového epitelu – crista ampullaris. Konce řasinek tohoto epitelu jsou vzájemně slepeny vrstvou gelu a tím jako celek registrují změny pohybu živočicha. Sluchový (akustický) orgán se vyvinul až u suchozemských živočichů. Sluchové buňky jsou uloženy v lageně, která se u obojživelníků protahuje, u plazů a ptáků tvoří dlouhý slepý kanál, který se u savců z prostorových důvodů spirálovitě stáčí v blanitý hlemýžď - ductus cochlearis. Od obojživelníků dochází k rozvoji středního ucha s 1 nebo 3 sluchovými kůstkami. U většiny obojživelníků a u všech plazů a ptáků je jedna sluchová kůstka – columella (kolumela). U savců se kolumela mění v třmínek – stapes, na který se napojuje kovadlinka – incus a kladívko – malleus. Kladívko naléhá na bubínek – membrana tympanica. U ryb dochází k vnímání zvuku pravděpodobně v sacculu. Dále je u ryb k vnímání (i tvorbě) zvuku využíván plynový měchýř, který je citlivý na chvění zvukových vibrací. Proto došlo u různých skupin ryb k vytvoření tzv. Weberova aparátu (systém modifikovaných částí prvních obratlů a žeber), který kaudálně nasedá na plynový měchýř a kraniálně na vnitřní ucho. K vývoji vnějšího ucha došlo až u některých plazů a ptáků a pak u savců.
U obratlovců došlo k vytvoření komorového oka – bulbus oculi. Na povrchu oční koule je bělima – sclera přecházející vpředu v průhlednou rohovku – cornea. Pod bělimou je cévnatka – chorioidea, která vpředu přechází v řasnaté těleso – corpus ciliare, na němž je zavěšena čočka – lens crystalina. Součástí střední vrstvy je i pigmentovaná duhovka – iris, která plní funkci clony před čočkou. Vnitřní stěnu zadní komory oční pokrývá sítnice – retina, v níž jsou uloženy světločivné buňky – tyčinky a čípky. Obratlovci s denní aktivitou mají převážně čípky (barevné vidění), zatímco v sítnici nočních obratlovců převládají tyčinky (vnímání světelné intenzity černobíle). U některých savců s noční aktivitou (šelmy aj.) došlo k vytvoření vrstvy buněk mezi sítnicí a cévnatkou (tapetum lucidum), která odráží světelné paprsky procházející sítnicí znovu na fotoreceptory a tím je zesílen zrakový vjem. Tento zpětný odraz se navenek projeví „svícením“očí. Jiní noční živočichové jsou vybaveni teleskopickým okem s konvexně vyklenutou rohovkou a válcovým bulbem (sovy) a někdy i přídatnou sítnicí (hlubinné ryby). K částečné nebo úplné redukci očí došlo u obratlovců žijících v trvale nepříznivých světelných podmínkách (hlubinné, jeskynní a podzemní druhy - některé ryby, z obojživelníků macaráti, ze savců např. slepci a krtci). Oči kostnatých ryb jsou zaostřeny na blízko a na dálku se zaostřují posunem kulovité čočky po optické ose k sítnici. U paryb, obojživelníků a amniot jsou oči zaostřeny na dálku a na blízko se zaostřují přitažením čočky k rohovce (žraloci, obojživelníci, hadi) nebo mění tvar čočky přitažením nebo uvolněním řasnatého tělesa (amniota s výjimkou hadů).
Dalšími smysly u obratlovců jsou chuť a čich. Chuťové buňky se nacházejí v chuťových papilách nebo pohárcích, které jsou obvykle na sliznici dutiny ústní, jazyka a hltanu. Čichový orgán – organum olfactus patří fylogeneticky k nejstarším smyslovým orgánům obratlovců. Čichové buňky jsou uloženy v párových (u bezčelistnatců v nepárových) čichových jamkách hlavy. Tyto jamky až druhotně získávají kontakt s dutinou ústní a tím i s dýchací soustavou prostřednictvím vnitřních nozder – choan (u některých svaloploutvých ryb, obojživelníků). U suchozemských obratlovců je čichový epitel uložen v zadní části nosní dutiny, kde pokrývá nosní skořepy. U některých obojživelníků, plazů a savců je vyvinut tzv. Jacobsonův orgán - organum vomeronasale. Jde o vychlípeninu stropu ústní dutiny situovanou na dno nosní dutiny, pomocí které lze vnímat i pachy, které přichází se slinami z dutiny ústní.
Trávicí soustava. Trávicí soustava začíná dutinou ústní – cavum oris, ve které bývá jazyk – lingua a často i zuby. Zuby mohou být morfologicky nerozlišené, tzv. homodontní nebo heterodontní, tj. morfologicky i funkčně různé. U fylogeneticky starších skupin obratlovců (ryby, obojživelníci, plazi) narůstají zuby nejen na čelistech, ale i na kostech patrového komplexu. Zuby difiodontní se obnovují jednou (mléčný a trvalý chrup), zuby polyfiodontní vícekrát. Na dutinu ústní navazuje hltan – pharynx, jícen – oesophagus, různě utvářený žaludek – ventriculus, syn. gaster, tenké střevo – intestinum tenue a tlusté střevo – intestinum crassum. Tlusté střevo je zakončené konečníkem – rectum, který vyúsťuje do kloaky - cloaca (např. obojživelníci, plazi, ptáci) nebo samostatným řitním otvorem – anus. Mezi tenkým a tlustým střevem mohou vyúsťovat 1-2 slepá střeva – cecum. Játra – hepar a slinivka břišní – pancreas ústí do přední časti tenkého střeva (duodenum). Vnitřní povrch střev je zvětšen pomocí střevních klků. U mihulí, paryb, svaloploutvých a paprskoploutvých s výjimkou kostnatých ryb je zvětšení střevní plochy vyřešeno spirální řasou (valvula spiralis, typhlosolis), která může probíhat až ke kloace.

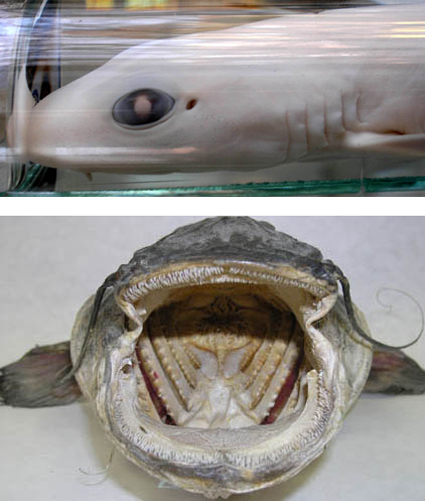
Obr. Významnou apomorfií (evoluční novinkou) obratlovců umožňující aktivní získávání potravy jsou čelisti vzniklé přeměnou žaberních oblouků. Obrázek schématicky ukazje původní stav u kruhoústých s devíti žaberními oblouky, které plní funkci opory žaberního aparátu. U paryb dochází k redukci prvních dvou oblouků a k přeměně třetího na čelisti. Zbývající žaberní oblouky mají funkci opornou. Z původních osmi žaberních štěrbin zůstává u paryb pět plus spirakulum - otvor za okem, který spojuje žaberní dutinu s vnějším prostředím. U savců žaberní oblouky zanikají, jejich rudimenty jsou zachovány jako ušní kůstky, části jazylky či chrupavky hrtanu (Obr. JK podle Ročka, 2002). Na snímcích hlava máčky (Scyliorhinus sp.) s dobře patrnými žaberními štěrbinami a spirakulem za okem a hlava sumce (Silurus glanis) se čtyřmi žaberními oblouky (Foto OS).
Dýchací a cévní soustava. Dýchacími orgány obratlovců jsou žábry nebo plíce. Někteří obratlovci jsou však schopni dýchat i kůží nebo ústní sliznicí. Žábry – branchiae mají pouze primárně vodní obratlovci. Pravděpodobně se jedná o modifikované filtrační zařízení, původně určené k zachycování potravy (pláštěnci, kopinatci, larvy mihulí), které se až později specializovalo i k dýchání. Žábry tvoří velký počet prokrvených lupínků v žaberních váčcích na přepážkách žaberních štěrbin (kterých je 4-15 párů), nebo přímo na žaberních obloucích. Žaberní štěrbiny ústí buď samostatně po stranách přední části těla, nebo do dýchacího prostoru krytého skřelemi. U bezblanných obratlovců, jejichž larvy (někdy i dospělci) žijí trvale ve vodě, se někdy vyvíjí přídatné dýchací orgány, umožňující často dýchat i atmosférický kyslík. Jde především o vnější žábry keříčkovitého typu (např. u larev bahníků, bichirů a obojživelníků). Dále to mohou být kostěné a vaskularizované labyrinty některých ryb (např. tzv. „labyrintky“- rájovci, čichavci) a plicní vaky u svaloploutvých (bahníci). Plíce – pulmo vznikly zdokonalením plicních vaků a jsou odvozeny od entodermu. Obojživelníci mají nejjednodušší vakovité plíce s nepatrně členitou stěnou, které připomínají plicní vaky dvojdyšných ryb. Teprve u amniot se zvětšuje respirační plocha a členitost plic.
S nutností dopravovat kyslík a živiny ke tkáním se zvyšujícím se metabolismem a zároveň odvádět od nich odpadní produkty látkové výměny souvisí rozvoj cévní soustavy. Tato soustava je tvořena sítí uzavřených cév- vasa (tepny – arteriae a žíly – venae). Srdce - cor má funkci svalové pumpy, která při smrštění (systola) vhání krev do tepen a při roztažení (diastola) nasává krev z žil. U primárně vodních obratlovců je žíla před vstupem do srdeční předsíně - atrium rozšířena v tzv. žilný splav – sinus venosus. Obdobně je rozšířena i část tepny vystupující ze srdce v tepenný násadec – bulbus arteriosus. Srdeční komora může být prodloužena v srdeční násadec – conus arteriosus. U ocasatých obojživelníků je přepážka v oblasti předsíně pouze naznačena, vyvinutá je u žab, ale komora je pouze jedna. První náznaky komorové přepážky se objevují u želv a u krokodýlů je přepážka téměř úplná. Úplně anatomicky i funkčně nezávislé komory jsou až u ptáků a savců. Z původně 6 párových oblouků aorty primárně vodních čelistnatců přetrvává u obojživelníků a plazů 4. oblouk jako levá a pravá aorta - z nich u ptáků zůstává jen pravá a u savců levá; perzistence pravého aortálního oblouku je u savců vývojová vada dlouhodobě neslučitelná se životem.

Obr. Evoluce srdce obratlovců: paprskoploutví; plazi – neúplná přepážka, pravý i levý aortální oblouk; ptáci - úplná přepážka, pravý aortální oblouk; savci – úplná přepážka, levý aortální oblouk. Aorta a její větve jsou znázorněny červeně, plicní arterie modře. (JK podle Millera a Harleye, 2005).
Krev (haema, sanguis) primárně vodních obratlovců je hypotonická vzhledem k mořské vodě, což ukazuje, že jejich počáteční vývoj patrně proběhl ve sladkých vodách. Výjimkou jsou sliznatky, které podobně jako mořští bezobratlí mají krev izotonickou s mořskou vodou, a paryby, kde se však jedná o odlišný mechanismus. Erytrocyty obratlovců jsou jaderné až na savce; ve fylogenezi se projevuje tendence k jejich zmenšování a ke zvětšování počtu.
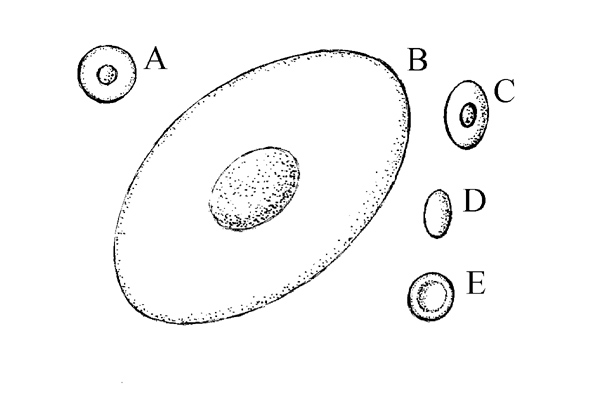
Obr. Erytrocyty obratlovců (jednotné měřítko): (A) mihule, (B) ocasatí obojživelníci, (C) paryby, paprskoploutví, žáby, plazi, ptáci, (D) velbloudi, (E) ostatní savci. (JK podle Gaislera a Zimy, 2007).

Obr. Protiproudový výměník tepla v noze arktického vlka a v noze kachny nocující na ledě. Uprostřed schéma přestupu tepla z arteriální krve (červeně) směřující na periferii do venózní krve (modře) vracející se do centra těla. To přispívá k zabránění úniku tepla do prostředí a udržení tepla v tepelném jádru těla. (JK podle Hickmana et al., 2004 a Millera a Harleye, 2005).
Vylučovací soustava. U obratlovců jsou vylučovacími orgány párové ledviny (renes) mezodermálního původu. Ledviny se zakládají v podobě segmentovaných nefrotomů uložených podél páteře. U mnohých zárodků a u larev mihulí se vylučovací ústrojí nazývá praledviny – holonefros (řec. holos = celý). Je uspořádáno segmentálně po celé délce trupu a jeho vývody ústí do primárního močovodu – Wolffův vývod. Z kraniální části holonefrosu se vyvíjí předledvina – pronefros, která zůstává funkční u mihulí, některých ryb a u larev obojživelníků. U holonefrosu a pronefrosu vzniká primární moč filtrací z klubíčka vlásečnic – glomerulus přes splanchnopleuru. U většiny ryb kraniální část vylučovacího orgánu tvoří pronefros a z kaudální části původního holonefrosu vzniká mezonefros – prvoledvina. U mezonefrosu dochází k vsunutí glomerulu do Wolffova vývodu, který dá základ pro vytvoření Bowmanova váčku, a tím vzniku nefronu. U dospělých obojživelníků je funkční pouze mezonefros. Jejich ledviny i ledviny mnohých dospělých vodních obratlovců (ryby aj.) jsou někdy označovány též jako opisthonefros, protože vznikají ze zadní části holonefrosu (řec. opisthen = zadní). U amniot vzniká definitivní (pravá) ledvina – metanefros se základní funkční jednotkou nefronem.
Rozmnožovací soustava. Obratlovci jsou odděleného pohlaví – gonochoristi (až na výjimky u ryb). Pohlavními žlázami – gonádami obratlovců jsou u samců varlata – testes a u samic vaječníky – ovaria. Původně se jedná o párové žlázy a jen ojediněle mohou být druhotně nepárové (častěji u samic, např. mihule, samice mnoha ptáků). Základem vývodných cest samčích pohlavních žláz je primární močovod (Wolffův vývod). Vývodem samičích gonád jsou vejcovody – oviducti vzniklé z Müllerovy chodby, která je embryonálně také odvozena od primárního močovodu. Distální část vejcovodů může být rozšířena v dělohu – uterus. Přídatné pohlavní žlázy jsou vyvinuty pouze u savců (s výjimkou Leydigovy žlázy žraloků). U ploutvovců a obojživelníků nejsou vyvinuty specifické pářící orgány, protože vypouští pohlavní produkty do vody a dochází k mimotělnímu oplození. U většiny plazů a ptáků probíhá kopulace přiložením nebo částečným vychlípením samčí kloaky do kloaky samice (vnitřní oplození). U hadů a ještěrů se vyvíjí k páření párová vychlípenina kloaky – hemipenis. U želv, krokodýlů a některých ptáků se můžeme setkat s náznaky pyje – penis, který se však v plné míře vyvíjí až u savců.
Určení pohlaví je u obratlovců překvapivě rozmanité. U bazálnějších skupin převládá environmentální způsob určení pohlaví, kdežto v následující fylogenezi se více prosadilo genetické určení, závislé na pohlavních chromosomech. U prvého typu (ZW - Abraxas), nazvaném podle rodu píďalek, je heterogametní pohlaví samičí, u druhého (XY - Drosophila) jsou heterogametní samci. U ryb, obojživelníků a plazů se vyskytují oba typy v pestré mozaice a pohlavní chromozomy mnohdy nebývají morfologicky zřetelně rozrůzněny. Oba typy mohou být přítomny i u různých populací téhož druhu. U ptáků jsou heterogametní samice (ZW/ZZ), u savců jsou heterogametní samci (XX/XY).